阅读:0
听报道

——基因组学研究追溯人类迁移历史(下)
编者按:
在过去十年,科学家对人类基因组数据的分析,正在重塑我们对人类演化历史及遗传适应的理解。2017年1月19日,来自美国、英国、丹麦和瑞典的六位学者在《自然》(Nature)发文,介绍了现代人及古人基因组测序分析为人类演化研究带来的一系列重大突破。
《知识分子》获《自然》与作者授权编译全文,介绍给关注人类命运与历史的中文读者。3月中旬,我们刊登了文章上半部分,今天介绍的是欧洲、亚洲、美洲和大洋洲人类的迁移史。
撰文 | Rasmus Nielsen, Joshua M. Akey, Mattias Jakobsson, Jonathan K. Pritchard, Sarah Tishkoff, Eske Willerslev
编译 | 汪鸿儒
审校 | 王传超
责编 | 陈晓雪
● ● ●
欧洲人起源
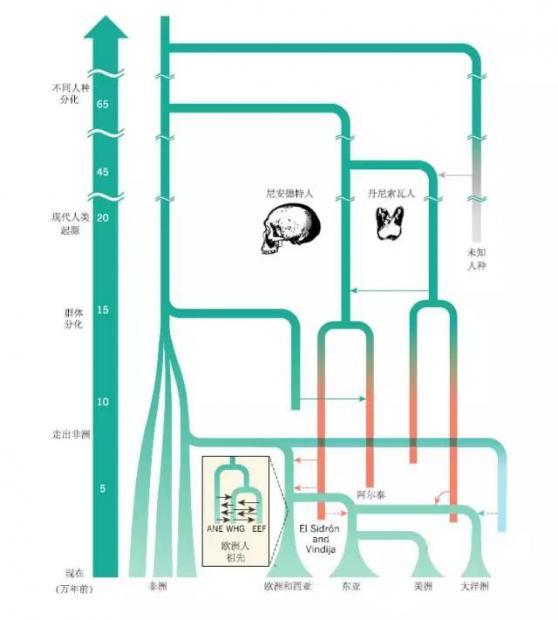

欧洲人群很可能含有三股或者更多的在不同时期涌入欧洲的遗传成分(图2和图3)。第一个现代人在43,000年前就出现在欧洲。这些旧石器时代的早期欧洲人,很可能对今天的欧洲人没有任何遗传贡献,因为证据显示欧洲人的遗传组成在末次盛冰期(译者注:是指末次冰期中气候最为寒冷、冰川规模达到最大的时期,距今约2.65至1.9~2万年)之前有过翻覆(译者注:在不同的时间,整个人群的遗传成分有着很大变动,是为翻覆),而这很可能和气候振荡有关。当然,早期欧洲人对现代欧洲人的遗传贡献具体有多少,仍有着争论。
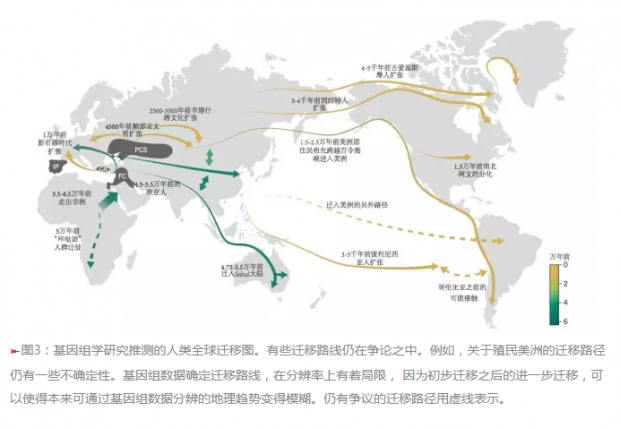
11,000年前左右,末次盛冰期过去以后,一种全新的基于畜牧和农业的新生活方式——也被称为新石器时代生活方式,出现在了新月沃地的多个区域(图3)。古DNA分析显示:这些地区的农民从安纳托利亚(Anatolia)中部地区扩张到了欧洲,但是新月沃地其他区域对早期欧洲农民遗传物质贡献有限。这些农民人群大致在七千年前到达伊比利亚半岛(Iberian Peninsula)(图3),并在大约六千年前到达大不列颠和斯堪的纳维亚(Scandinavia)。
新石器时代人遗存的基因组数据分析表明,农业文明自新月沃地向欧洲传播,由农民人群的大规模迁徙所驱动的,并对本地的采集狩猎人群进行同化吸收,这证明了新石器时代生活方式的传播是通过人群的迁徙实现的,而不仅仅是思想或者文化的传播。根据从基因组数据估计出有效群体的大小推测,新石器时代的生活方式帮助增加了区域人群的规模。但考古学的数据显示,这些农民的健康状况有时很不好,因为(他们的遗骨)有着大量营养不良和骨溃疡的痕迹。
在新石器时代晚期和青铜时代早期,另外一波进入欧洲的迁徙为欧洲引入了第三股遗传成分。4,500年前,来自颜那亚文明(Yamnaya Culture)东欧大草原的牧人参与了进入中欧的迁徙。这些牧人自己又是俄罗斯和高加索地区多个狩猎采集人群的后代。这次迁徙很可能与征服以及技术创新有关,比如骑马;迁徙可能把印欧语传播到了欧洲,虽然有一些语言学研究者认为新石器时代的欧洲农民已经说印欧语了。新石器时代晚期和青铜时代无疑是一个动荡的时期,其间,来自大草原的牧人将其遗传物质扩散到整个西欧和北欧。
当代欧洲人群的三股遗传成分反映了:
(1)末次盛冰期结束后,采集狩猎人群在欧洲的扩散繁衍;
(2)安纳托利亚农民向欧洲迁移;
(3)新石器时代晚期和青铜时代来自东方的人群迁移。
这些成分在很大程度上可以解释当代欧洲的遗传多样性。例如,新石器时代的遗传成分似乎在南欧人群,如撒丁岛人中最多。当代欧洲人中的遗传变异和地理有着强烈的相关性,并且越向北,多态性越低。尽管欧洲主要的遗传成分由几波独立的迁徙引入,但是后续受地理限制的基因流动塑造了现在的欧洲人遗传变异。因此,在史前欧洲,相较于地理因素,文化和生活方式是决定基因组异同更为重要的因素。
亚洲人的起源
目前研究的大多证据显示,亚洲至少有着两波早期的迁徙事件。第一波包含澳大利亚和巴布亚人的祖先,另外一波包含东亚人的其他祖先,这两波迁移之间存在基因交流(图2和图3)。还有一些证据暗示亚洲只有一次迁移事件。然而,亚洲人祖先最早具体是如何迁移扩散开来的,基本上还是未知。
目前只有两个早期东亚人的基因组被测序。一个基因组来自于西伯利亚中南部马耳他-布里特(Mal’ta-Buret’)文化、生活在24,000年前的一个个体(MA1)。这一个体与西欧人以及美洲原住民有着很强的遗传亲缘性,但与东亚以及西伯利亚人亲缘关系较弱,这意味着旧石器时代晚期遗传信号的地理分布和现在有着很大的不同。另外一个基因组来自西伯利亚乌斯季伊希姆(Ust’-Ishim)地区一个45,000年前的个体。当把丹尼索瓦人遗传渗入考虑在内时,这一个体显示出与西欧、东亚及澳大利亚原住民人群相同的遗传亲缘性。连同一个来自于欧陆俄罗斯(译者注:俄罗斯的欧洲部分)36,000-38,000年前的个体(Kostenki 14, K14)基因组证据,分析表明,K14 与当代西欧人亲缘关系很近,但与东亚人并不近,据此推测,西欧和东亚人的分化发生在36,000-45,000年前左右(译者注:乌斯季伊希姆的研究结果暗示45,000年前欧洲人和亚洲人还没有分化;而K14的研究结果暗示36,000-38,000年前,欧洲人和亚洲人已经分化)。
一份包含青铜时代欧亚大陆101个古人的低深度测序基因组研究显示,后来有两次从西亚和欧洲向中亚的人群扩张导致了亚洲人群的遗传混合,或者局部替代了和MA1相关的采集狩猎者。第一次扩张是颜那亚牧人在大约5,000年前进入亚洲,这与颜那亚人群扩张进入欧洲发生在同一时代。随后,在2,500-3,500年前,中亚的颜那亚人(形成了阿凡纳羡沃文化,Afanasievo Culture)被来自于辛塔什塔文化(Sintashta Culture)的人群所替代了;这些辛塔什塔人由乌拉尔和欧洲迁入,并且和东亚人发生了混血。
大洋洲人的起源
考古证据显示,47,500-55,000年前,现代人就出现在大洋洲。古人头骨形态变异数据,支持至少有两次独立的人群迁移进入古老的Sahul大陆——包含现在的澳大利亚、新几内亚和塔斯马尼亚。语言学的数据、石器制造工艺以及驯化物种引入(如澳洲野犬)的证据,也支持类似的观点。
然而,目前唯一一个对澳洲土著和巴布亚人的群体基因组学的研究,支持整个Sahul大陆人群主要来源于一波迁入事件,随后巴布亚人和澳洲土著的祖先人群发生了分化。而澳洲土著人群又进一步发生遗传分化,这可能与环境的变化,如沙漠化有关。因此,直到近代,澳洲土著人似乎一直居住在一个高度隔绝的环境中。
波利尼西亚人分布在一个三角形的南太平洋岛屿群上,东部以复活节岛(Rapa Nui)为界。一份基于大洋洲的现代人全基因组数据的研究证实了考古学关于波利尼西亚人的推测:波利尼西亚人有着美拉尼西亚和东亚人的混合血统。最初的波利尼西亚人扩张中,主要为原始的东亚血统,美拉尼西亚血统是后来才加入的。波利尼西亚人向东扩张结束于大约1000年前,在扩张的过程中,他们是否到达过美洲并且与美洲土著人发生过混血,仍有争议。一份对古代南美洲鸡遗存的遗传学研究支持发生过混血,但这份研究也受到质疑(译者注:2007年一份研究报告中,奥克兰大学的研究者称在智利发现了早于1492年,即哥伦比亚发现新大陆之前的鸡骨头,DNA研究也显示它和波利尼西亚早期的鸡有着亲缘关系,因此暗示波利尼西亚人和南美洲人有着早期的交流。但2008年,来自澳大利亚的研究人员通过分析智利本地大量鸡的样本及全球1000余份鸡DNA数据,指出2007年研究中的鸡的样品应起源于欧亚,因而不支持波利尼西亚人和南美洲人有着早期的交流)。
对一个1650年左右的巴西人基因组测序研究发现,这个个体与现代波利尼西亚人有着很强的亲缘关系,而1650年又早于波利尼西亚奴隶进入南美的记载。一方面,这份研究为波利尼西亚人和美洲土著人的早期接触提供了证据。但另一方面,这也可以解释为更早的波利尼西亚奴隶进入南美,但是未被记录。目前一个更有说服力的结果是,现代复活节岛居民全基因组的统计研究,支持了复活节岛人群基因组,在1280-1495年左右,曾和美洲原住民有着遗传混合。而这个时间比欧洲人最早在1722到达复活节岛要早几百年。然而,要最终解决是否有早期接触的争论,得看是否能够在早于1492年的人遗存中发现波利尼西亚和美洲土著混血的证据。
美洲人的起源
现代人类最早出现在美洲大陆的证据要追溯到大约14,000-15,000年前,而美洲广泛的殖民则随着克洛维斯文化(Clovis Culture)的出现而开始。克洛维斯文化也是美洲最早、研究确切的考古文化。
然而,直到13,000年前左右,北美大部分地区仍被巨大的冰盖覆盖,这使得人们从白令陆桥(现在的西伯利亚东北部和北美的西北部)进入美洲南部变得异常困难。融冰之后,一个长约1500千米、无冰雪的内陆走廊就形成了。对加拿大湖芯的宏基因组分析(译者注:对环境样品直接进行基因组测序并分析),估算出这一走廊在12,600年前左右才有生命迹象。因此,这个走廊不大可能是克洛维斯人和之前人群南向迁徙的早期路径,尽管也有一个关于北美野牛的研究与这一结果不一致。美洲人的早期祖先,是如何以及何时跨越更新世的冰盖进入北美南部的,我们并不清楚。我们也不知道克洛维斯人和之前人群的迁移是否属于同一次迁移。最可能的情形是,大约在14,000年前,美洲人祖先沿着北美西海岸南向迁移,随后也不断向内陆渗透(图3)。
基于头骨形态和石器的分析,曾有人提出早期美洲人不是当代美洲原住民的直接祖先,而与澳大利亚-美拉尼西亚人、波利尼西亚人、日本的阿伊努人等其他人群有关联。然而,基因组学的研究在很大程度上否定了这些模型。
2014年,丹麦哥本哈根大学研究者主导的一项研究发表了最古老而且唯一与克洛维斯相关的人(发现于美国蒙大拿)的基因组,这一个体生活在距今12,600年前。分析指出,克洛维斯人群是很多当代美洲土著人的直接祖先。类似地,发现于美国的华盛顿州的9,500岁的肯纳威克人(Kennewick Man),起初因为头骨形态被认为和阿伊努人及波利尼西亚人紧密相关。然而基因组序列的分析判定,这个肯纳威克人和当代的美洲原住民的关系最为密切。此外,早期进入美洲的迁徙孑遗人群(relicts)起初被认为和澳大利亚–美拉尼西亚人紧密相关,后来也被证明遗传上和当代美洲原住民相近。
基于全基因组序列,来自丹麦哥本哈根大学和加州伯克利大学等机构的研究者估计美洲原住民基因库形成的时间应早在23,000年前。这又为美洲人祖先较早进入美洲提供了进一步的证据。但把美洲最早的考古遗址年代考虑进来,美洲人的祖先和他的西伯利亚的祖先分开后,在向东迁入美洲以前,可能一直在西伯利亚或者白令陆桥处在隔绝状态直到8000年以后。在美洲以外,虽然现代西伯利亚人是美洲原住民最近的“亲戚”,但对24,000年前的马耳他人(MA1)骨架的基因组测序分析指出,美洲人来源于马耳他人和其它一个或者多个未知的东亚人群混合的群体。由于克洛维斯人相关的基因组和当代美洲原住民含有同样多的马耳他人遗传印迹(14–38%),因此,推测混血事件发生的时间应早于12600年以前。不过,这些遗传混合发生在美洲之内还是之外仍然不清楚。
美洲原住民的基因组数据曾被用来确认一起早期群体分化事件的地理位置——这一事件发生在大约14000-13000年前。分化形成的南部分支包括说美洲印第安语(Amerindian-language)的人群,而北方的分支包含说阿萨巴斯卡语(Athabascan-language)以及其它语言,如克里语(Cree)和阿冈昆语(Algonquin)的人群。基于全基因组测序数据分析对分化时间的估计显示,两大分支人群同时从西伯利亚人群分化出来,这意味着美洲印第安和阿萨巴斯卡人群来源于共同的祖先人群,之后,他们接受了来自亚洲的基因流。美洲原住民这两大分支的分化,是发生在西伯利亚还是美洲冰盖的南部或者北部仍在争论之中,需要更多的古基因组研究。类似地,当代巴西美洲原住民中发现的澳大利亚-美拉尼西亚的信号(图3)是由于后来的基因流,还是一个未知的早期迁入的人群造成的,仍也未确定。到目前为止,还没有在任何美洲的古人基因组上发现这一遗传信号。
北美靠近北极圈的因纽特人(译者注:即爱基斯摩人,因“爱基斯摩”的称呼带歧视意味,现已改称)已被证实起源于独立于美洲原住民的迁移事件。然而,对于出现在5000年前的美洲,如今绝迹的古爱斯基摩文化,他们到底代表了现今因纽特人的祖先,抑或代表了独立的另一支来自于西伯利亚人群?这一问题也存在争论。通过对格陵兰的一簇4000年前的头发DNA测序,科学家们发现头发的主人通过一次独立于美洲原住民与因纽特人的事件,从西伯利亚迁徙而来(图3)。这一人群凭借其独特的生存策略与技术,在北极生存了大约4000年,但最终在700年前左右被因纽特人取代。
人类对新环境的适应
遗传数据的分析不仅告诉我们人类的演化历史,还告诉我们自然选择是如何影响人类这个物种的。人类的扩散首先发生在非洲,随后到全球其他地区,他们遇到新的环境条件带来了选择的力量,这包括冰河时代美洲和欧洲大部的严寒,新遇到的病原菌和光照条件。文化上的创新,如改良的打猎和捕鱼方法、动植物的驯化,以及饮食的变化,也带来了新的环境条件。基因组测序数据让我们有机会去系统扫描人类基因组上受选择的区域。通过利用大量可用于人类基因组分析的资源,我们有可能确定每个受到选择的遗传变异的具体的功能。因此,DNA序列分析发现的选择证据与这些序列在适应局部环境中扮演的角色之间建立了一种联系。此外,古DNA的研究使得我们可以直接观察等位基因频率随着时间的变化。
对局部环境的适应
当人类走出非洲,他们遇到的一个最明显的改变是,在高纬度,阳光照射减少了。居住在赤道附近的人群有着深色的皮肤,这保护人们免受紫外线辐射造成的皮肤损伤和叶酸降解。然而,紫外线辐射对维他命D的产生也有重要的催化作用,而维他命D又是骨骼发育和健康所必须的。居住在高纬度的人群接受紫外线辐射剂量较低,他们的淡色皮肤被认为能够更高效促进维他命D的合成。在高纬度地区,偏好少量皮肤色素的自然选择影响了多个基因,包括MC1R, SLC24A5和SLC45A2(也叫作 MATP)。
人类适应局部环境的另外一个典型例子出现在生活在高海拔地区低氧环境(如西藏)的人群。研究显示,适应了高海拔生活的藏人对低氧环境有着不同的红细胞生成调控机制。基因组研究揭示这种适应是由低氧应答通路中的两个基因,EPAS1和EGLN1的等位基因频率变化所驱动的。饮食上的改变,特别是那些和新打猎技术或耕作方式出现相关的改变,也对人类基因组有着相当大的影响。最著名的例子是乳糖耐受性的自然选择,它影响了欧洲和非洲乳品业人口的LCT基因的调控。同样地,FADS家族的基因,它们的产物催化多不饱和脂肪酸的合成,似乎也在几次转向或转出素食的生活方式转变过程中受到选择。
然而,最重要的局部适应的驱动因素,很可能是局部的病原菌环境。具体的例子包括编码主要组织相容性复合体(MHC)的基因,因为MHC将外源的多肽呈递给免疫细胞;beta球蛋白基因位点;以及非洲人的G6PD基因,在这个基因上特定突变的杂合体携带者会有登革热的抗性。
我们从人类的自然选择研究中学到……
从人类选择的全基因组研究可以汲取几个重要的经验教训。首先,大多最初的基因组选择扫描研究提出,选择会即刻作用于新突变,但现在看来,在许多例子中,选择往往作用于已有的变异——也就是,等位基因在被选择之前已经存在过一段时间了。此外,这些变异在很多情况下是通过和其它人种发生混血而渗入的。越来越多从尼安德特人或者丹尼索瓦人渗入现代人类的遗传变异被鉴定出来。例如,藏族人中的适应性的EPAS1单倍型似乎就来自丹尼索瓦人,而且对主要组织相容性复合体基因(MHC)和MC1R基因的选择都很可能是通过来自于尼安德特人的基因渗入实现的。人类走出非洲并遇到新的环境,与其他已经适应这些环境的人种进行混血似乎是快速适应新环境的一个重要因素。这可能尤其适用于和免疫及防御感染相关的基因,因为现代人类很可能遭遇到来自于其它人种的病原体,而人类对此还没有免疫力。
其次,许多影响了人类基因组的选择是为了应对人类自己引起的环境变化,如文化创新驱动的饮食上的改变,社会结构的改变、城市的出现引起的人群病原体负荷增加。当我们改变周围的环境,产生的条件改变又引起新的选择压力。因此,生物演化和文化演化是紧密联系在一起的。
第三,受到选择的遗传变异和对人类健康有着很大影响的遗传变异往往关系紧密。人类演化的研究因此和医学遗传学有着越来越强的关联。例如,适应高海拔而受到选择的变异为研究高血压提供了一个模型。类似地,已被证实受到选择的和饮食适应相关的变异,如FADS基因家族的基因,可辅助开发基于基因组的个性化饮食。
第四,在不同大陆人群等位基因频率差别最大的基因往往和可见的性状,如皮肤、头发和眼睛的色素有关。一个有趣的结果是,从色素的角度来说,不同的地理人群的差异比基因组实际平均水平“看”起来更加地不同。因此实际上,来自不同地区的人,在遗传上,比他们的发色、肤色以及其他可见性状看起来更加相似。
作者介绍:
Rasmus Nielsen, 美国加州大学伯克利分校计算生物学教授
Joshua M. Akey,美国华盛顿大学基因组学系教授
Mattias Jakobsson, 瑞典乌普萨拉大学有机生物学系教授
Jonathan K. Pritchard,美国斯坦福大学遗传学系与生物学系教授、霍华德•休斯研究员
Sarah Tishkoff,美国宾夕法尼亚大学遗传学系与生物学系教授
Eske Willerslev,丹麦哥本哈根大学地质遗传学中心主任,英国剑桥大学动物学系讲席教授,英国威康信托基金会桑格研究所副研究员
编译:汪鸿儒,遗传学博士,中科院古脊椎动物与古人类研究所古DNA实验室博士后
审校:王传超, 人类学博士, 美国哈佛大学医学院遗传学系、德国马普人类历史科学研究所博士后
原文标题“Tracing the peopling of the world through genomics”,2017年1月19日发表于《自然》(Nature)杂志。《知识分子》获作者与《自然》授权编译。
原文链接:

话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}