撰文|王家槐(北京大学客座教授)
责编|李晓明
那是2013年北京一个典型的仲夏之夜。白天的酷热已然散去,清凉的晚风习习。走在花园般美丽的北大校园,树荫底下灯影斑驳,芬芳之味令人陶醉。我还在回味刚刚在漂亮的百年讲堂看的一场电影,突然看到手机里有好几个未接电话。正要查,电话铃又响了。那是我的博士后,Lorenzo Finci。他无比兴奋地告诉我,我们的课题取得重大突破!他是指我们正在研究的对中枢神经系统发育起关键作用的轴突导向蛋白的分子结构。
事情还得从四年前我去拜访饶毅在北京的家说起。我1990年左右在哈佛就认得饶毅了。那时他是知名干细胞生物学家Doug Melton的博士后,我在同一个系里做访问学者。大概在2009年左右,我风闻饶毅要辞掉在美国西北大学讲席教授的职务,到北大生命科学院当院长。我决定利用回国的机会到他家聊聊。不料我们的会面竟变成了三个半小时的学术沙龙。饶毅给我滔滔不绝地介绍起令人入迷的神经发育机理,他还随手从茶几上拿起一张小纸,在上面涂来涂去,给我这个门外汉描画起神经是如何生长的。我知道这是他在美国做出过不小贡献的领域,此情至深。他的结论是,“从神经生物学角度来看,问题已经都清楚了。”“那么它们的分子机制呢?”我反问了一句。饶毅似乎没有思想准备,顿了一下,才说“那倒好像还有很多可探索之处。”说完沉思了一会儿。然后,他突然站起来问我“你有兴趣到北大来研究这个问题吗?”他见我没有吭声,又以一种十分肯定的语气对我说,“你很快会看到,在中国做研究会是一个很好的选择。”

那次聊天以后,饶毅又动员了我好多次到北大,还真使我动心了。我研究细胞表面受体的分子结构已有很多年,主要对象是免疫受体。受体分子一般由一个一个小的模块堆积而成,犹如是一颗一颗珍珠穿在线上。细胞用这种从表面伸出来的受体来感受外部环境,进而做出相应的反应。从分子结构角度来看,免疫受体和神经受体没有很大的差别,只是它们的生物学功能不同而已。我完全可以用我已有的免疫受体结构的知识去探索一个新的领域。后来,我曾经在清华和施一公开玩笑说,我们搞结构的可以摇身一变,从研究免疫变成研究神经啦。说实话,我从年轻时就对北大很向往,在科学院生物物理所工作时也和不少北大的朋友一起有过愉快的合作。我也特别喜欢和高校中朝气蓬勃,二十多岁的年轻学生在一起。经得我们医学院的同意,我决定到北大试试。在当时教育部的985经费的支持下,我在北大生科院里组建了一个小组,有两个博士后,一个技术员,一个从科大招来的研究生,一个本科生和一个秘书(图1)。我们主攻神经生物相关的结构研究,也和院内其他老师合作开展药物作用的分子机理等。我也非常高兴地承担了相当的教学任务。

Lorenzo是我在北大的第一个博士后。2011年秋天他已经在波士顿大学完成了博士论文答辩。他看到了我发出的招聘广告。在仔细阅读了我以前发表的文章和我打算在北大开展的课题后,他产生了极大的兴趣,决定到我在波士顿的办公室来面谈。我们谈得很投机。他显然是一个很有抱负的年轻人。我当时告诉他,跟我去北京,要有几个思想准备。第一,我无论怎么努力争取,也只能付你相当于在美国当博士后一半的工资。第二,我在北大是要新建一个实验室,条件和我在波士顿的实验室有不小的差别。第三,我在北大的时间一年里不会超过半年。第四,北大大概还只有极少极少外籍博士后。基于这四点,我告诉他,你要有好好适应的思想准备。他说这课题太有吸引力了,他还是决定参加我在北大的研究小组。钱多钱少他不在乎。反正现在还不是挣钱的时候。他还笑着说,“我就是要做一个最早从美国到中国做博士后的学生之一。”没过几个星期以后,他兴冲冲的又来找我,递给我刚刚拿到的博士毕业证书,握住我的手说,带我去北京吧!我拿着那似乎还有油墨味的证书,很是感动。经过在我波士顿的实验室里熟悉了三个月后,一点中文都不懂的Lorenzo就这样于2012年刚过元旦就飞到了北京。我比他早两天到京,我和实验室先期入组的两个女孩去机场接他。他此前从未坐过这么长时间的飞机。当他推了行李车出来时,脸色煞白,有气无力地对我说,“家槐,我真累坏了。”这把我吓了一跳。到底还是年轻,等当天下午,我们把他安顿到专门为他申请到的漂亮的留学生单元宿舍后,他已恢复生龙活虎,高兴得不得了。我为他配了一个基本能用英语交流的技术员。他们很快就把工作开展了起来。
我们要研究的神经受体叫DCC。DCC最初被发现时是结肠癌细胞的标记受体,后来才证实,它其实更重要的是神经元细胞表面的受体。在神经系统早期发育阶段,让各个神经元细胞正确地互相连接起来,建立神经网络,是极端重要的。在这个过程中,每个神经元细胞会伸出一个叫做轴突的部位,它会在一个叫Netrin-1(神经生长因子)的信号分子以及其它一些分子引导下生长,一直抵达目的地,与它应该连接的神经元细胞建立突触。Netrin-1是第一个被发现的引导轴突生长的蛋白化学因子。它还在其它很多细胞功能中发挥作用。DCC是Netrin-1的主要受体。正是Netrin-1和轴突表面DCC的相互作用,引导了神经元的轴突向正确的方向延伸。在2011年,关于Netrin-1分子的结构以及它如何与DCC相互作用的机理,这个神经发育领域多年来探讨的问题,基本上还是一个未知数。Lorenzo和他的合作伙伴张婕的课题就是要负责制备DCC和Netrin-1,把它们按照一定比例混合在一起培养成晶体,然后用X-射线晶体学的方法来测定其分子三维结构。我们的兴奋点是想看看Netrin-1是如何与DCC相互作用,从而使神经元细胞做出相应的反应。
这是一个多年来在国际上竞争非常激烈的课题。谁都想知道,Netrin-1和DCC到底是如何工作的。这毕竟是神经生长领域里人们最关心的问题之一。据我们所知,当时牛津,哥伦比亚和大阪大学都有人在做。我早年的一个博士后Rob Meijers在德国汉堡也在尝试,而且他在制备Netrin-1上已经有很好的突破。我马上和Rob联系,建议我们分工合作,因为在Lorenzo抵京前,张婕在DCC制备上也已有进展。为了保证合作可以愉快、顺利进行,我和Rob明确提议,将来如果我们的合作成功,他可以是文章的最后作者,Lorenzo署名第一,北大将是第一单位,我做一个共同通讯作者就可以了。就这样,我们在2012年春天顺利达成协议。双方全力以赴,集中精力攻科学难关。Lorenzo 和张婕配合得非常好,解决了很多技术问题。为了提取高质量的蛋白,他们常常加班。Rob在汉堡的实验室就位于世界上最好的同步辐射X光光源之一。理想的安排是我们把DCC蛋白送到汉堡,和Rob的Netrin-1混合后培养出DCC/Netrin-1二元复合物晶体,然后直接在汉堡收集X光衍射数据。当时在中国和德国之间没有好的快递业务。为了保证娇贵的蛋白在运输途中不变质,可怜的Lorenzo 只好多次往返于北京和汉堡之间,用冰盒运送样品。少不了会在中国和德国海关有很多周折。很快,也就一年半左右,在双方共同艰苦努力下,在2013年六月Rob好不容易获得了三颗晶体,只有三颗,而且很小,挺不好看,一颗还不幸坏掉了!Rob小心翼翼取了其中剩下的一颗晶体,安到了衍射仪器上。哇,没想到,竟然有三埃左右的衍射分辨率!作为一个训练有素的晶体学家,只要收到了合用的数据,解析这个结构,并没有费Rob很大的功夫。这就是他迫不及待把这个激动人心的消息用电子邮件迅速发给在北京的我们的那个仲夏之夜。
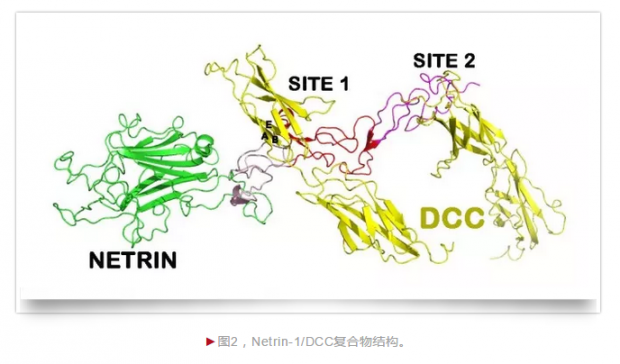
我们怀着极大的兴趣开始分析这个结构。一个没有想到的发现是,一个Netrin-1在两个不同的位点各结合了一个DCC(图2)! 我和Rob以及Lorenzo对此展开了激烈的争论。我认为后来被称为结合位点1是对的,符合一般的蛋白质相互作用的规律,而另外一个位点是假象。Rob认为那个位点2是对的,更能和生物学数据对得上。Lorenzo 则笑着说,如果家槐对,我们喝青岛啤酒庆祝;如果Rob对,我们喝Heineken(全世界有名的荷兰啤酒,因为Rob本是荷兰人。在Rob的生日那天,我们寄给他的礼物是这张图3)。

经过仔细阅读文献,原来神经生物学家已有文章报道,如果Netrin-1结合了两个DCC受体,轴突的反应是一种趋向性生长;如果Netrin-1 结合一个DCC,同时又结合另外一个受体叫UNC5, 那么轴突的反应是背离性生长。也就是说我们观察到的两个位点可能都对。仔细分析后的推断是,位点1是专门结合DCC的,位点2可以结合DCC,也可以结合UNC5。这样我们有可能找到了Netrin-1生物学功能的分子机理。这时,我们三个人兴奋极了。我那几天正好在北京。我把饶毅找来看我们的结果。他也很兴奋,和我一起找到了他推荐给我的在北大的研究神经生物学的合作者,张研老师。在讨论中,我请张老师根据我们的推测,做神经生物学实验来验证。现代结构生物学需要的就是结构和生物功能的紧密合作。张老师实验室花了不少时间,作了非常严谨的工作。她们的结果很漂亮地证明了我们关于两个位点的生物学功能的假设。这项研究于2014年在神经生物学领域内最热门的杂志Neuron上发表。
科学的发展常常会曲折前进,难以预测。当年饶毅对我说的,关于轴突导向,“从神经生物学角度来看,问题已经都清楚了。”到了2017年,问题突然又不那么清楚了。国际主流神经生物学界对于建立神经网络有了新的看法。经典的轴突导向理论认为,轴突表面上的DCC受体是感受着Netrin-1在环境中的浓度梯度前进的,是一种趋化性。但是很久以来一直不是很明白,生物体内这个Netrin-1的浓度梯度究竟是如何建立起来的。2017年出现两篇重磅文章,分别发表在Nature和Neuron。作者们认为,轴突的导向并不一定是来自于长距离Netrin-1 浓度差引起的“趋化性”,更可能是在轴突前进的路上,沿途产生的Netrin-1与轴突表面的DCC在短距离内发生了作用。是一种“趋触性”。形象点说,轴突不是游泳前进,而是爬行前进,有点像常春藤在墙上爬。这涉及到细胞和细胞之间的触碰。说来有趣,我们和Rob刚刚合作完成的另外一个课题,正赶上为此提供了一个非常及时的分子结构证据!
我在北大的研究小组总的方向是轴突导向的分子机理。和Netrin-1/DCC课题一起开始做的还有另外一个导向分子,叫Draxin。那是2009年由一个日本研究小组新发现的蛋白分子,也作用于DCC。与Netrin-1不同,Draxin和DCC的作用只会引起轴突的背离排斥作用。 与Netrin-1另外一个不同之处是,Draxin似乎是个冷门。当时不少主流神经生物学家好像都对它不太感兴趣。那个课题是由我在北大的另外一个博士后刘迎负责的。刘迎做得很辛苦。因为要用比较复杂的真核细胞来表达这个糖蛋白。实验要求比较苛刻。而且她一个人要同时做Draxin以及DCC分子与Draxin相互作用的那个部位。经过两年多的努力, 几经周折,大概在Lorenzo的文章于2014年发表的前后,刘迎也拿到了晶体。但是,令人困惑的是,晶体的样子和我们几年前在波士顿得到的只有DCC那个部位的晶体非常像。也就是说刘迎得到也许不是Draxin和DCC的复合物晶体,而是我们已经发表了的单独只含DCC的晶体。更困惑 的是,当我们在日本的同步辐射实验室收数据时发现,刘迎的晶体居然和单独DCC的晶体有完全一样的物理参数。当时我也在现场。可以想象,如果真的不是复合物晶体,那会多么令人沮丧!那毕竟是刘迎两年多的辛勤劳动!蛋白质晶体有一个特点。蛋白质分子实际只占晶体大概一半的体积,其余空间填满了溶剂分子。根据计算,刘迎那个晶体,溶剂含量尤其高。因此,我们还抱有一定的希望,也许Draxin 还是有可能在里面。我们在现场很快用收到的数据粗粗作了一些计算,似乎在DCC分子的一个部位看到一点别的东西。回到北京后,经过我们另外一个合作者,一个非常有经验的晶体学家肖俊宇老师的仔细分析计算,最终确定,原来Draxin只用了它大约五分之一的部位和DCC作用,其余部分在晶体空间中处于无序状态。刘迎那个高兴真正是很难用语言来形容的!肖老师后来在上海新的同步辐射光源又收了一套分辨率更高,质量更好的数据。结构精度有很大的提高。就在这时候,Rob的实验室也解析了一个结构,那是Draxin的另外一个很小的片段与Netrin-1相互作用的复合物。也就是说,Draxin既能与DCC结合,又可以和Netrin-1在另外一个部位相互作用。
在获得了Draxin/DCC和Draxin/Netrin-1这两个结构以后,我们一直在思索它们的生物学意义。 与此同时,张研老师的实验室也作了很大的努力。她还派她的得力学生刘一穹到汉堡去作实验,从神经生物学的角度来探索两个结构的功能。Rob两次来美国, 到我在波士顿的办公室作深层次的研讨。波士顿,北京和汉堡之间的视频电话更是不计其数。当我们看到2017年那两篇文章后,顿时给了我们很大的启发。我们意识到,如果把两个结构合在一起,我们可以提出一个很诱人的假设。因为既然我们已经看到了 DCC/Draxin,Draxin/Netrin-1和Netrin-1/DCC一系列的相互作用,我们可以设想在两个神经元的轴突之间可能有一个由DCC-Draxin-Netrin-1-DCC连起来的桥梁。那不就是细胞和细胞之间的接触吗!我们以此思路改写先前的稿子,还是投到Neuron。2018年元旦刚过没几天,文章就被接受。
还是那个2013年北京的仲夏之夜,当我和Lorenzo 肯定我们的课题有了突破,兴奋之余,Lorenzo 提议去喝一口。这个美国小伙子在北京一年多生活下来已经比我这个原来的老北京对北京还要熟悉多了。他说白石桥那儿的西苑饭店里有一个理想之处。我们当即打个出租前往。那是一个不小的露天酒吧,中式的庭院风光,却兼有西式的情趣。月光下,夜色正美,端的是一个好去处。反正Rob又不在,我就要了两瓶青岛啤酒,我们两人就着花生等干果,边喝边讨论下一步计划,真是其乐融融。而2018年初当Rob知道Neuron接受我们的第二篇文章后,他说他当晚将用法国的苹果白兰地来庆祝。我告诉他, 我极不善饮,但是我很喜欢有点甜味的法国的橘子白兰地。是我的家藏之一。今晚我将也来一呡。张研老师则在实验室开了香槟。 当我用电子邮件联系刘迎时,问她如何庆祝。她说,王老师,要不你和已经回美国的Lorenzo,以及你把她推荐到英国留学的张婕,你们啥时来北京,咱们一起也找个好去处去畅饮?
作者注:感谢华庆新和李小龙阅读并提出很好的意见,修改本文。
参考文献
1、Finci LI, et al. The Crystal Structure of Netrin-1 in Complex with DCC Reveals the Bifunctionality of Netrin-1 As a Guidance Cue. Neuron. 2014. Doi:10.1016/j.neuron.2014.07.010.
2、Dominici C et al. Floor-plate-derived netrin-1 is dispensable for commissural axon guidance. Nature. 2017. doi:10.1038/nature22331.
3、Varadarajan et al., 2017, Neuron 94, 790–799.doi: 10.1016/j.neuron
饶毅回忆
▼▼▼
每个人对事情的记忆和理解不同。我实验室从1999年开始研究排斥性导向分子Slit及其受体和信号转导通路,而2004年开始研究netrin的信号转导,到2009年在PNAS发的DSCAM,已经使我认为需要做结构生物学了,要不然不好解释机理。我在这之前鼓励过北大苏晓东老师做,原因是netrin的神经生物学比较清楚,但它的分子机理不清楚,早期既能结合吸引性受体DCC、又能结合排斥性受体UNC5,而后来又发现介导netrin吸引性的还有DSCAM。这样就很奇怪,用神经来做研究显然不够,而结构的研究可能有帮助。所以,我应该是认为这是结构生物学研究的好问题,而王老师是结构的专家。我是1991年到哈佛的生物化学与分子生物学系做博士后的时候认识王老师的,那时王老师已经在哈佛发过多篇结构生物学的Nature文章。
文献:
1、Liu G, Beggs H, Jürgensen C, Park HT, Tang H, Gorski J, Jones KR, Reichardt LF, Wu JY, and Rao Y (2004). Netrin requires the focal adhesion kinase and the Src family kinases to induce axon outgrowth and to attract axons. Nature Neurosci 7:1222-1232.
2、Liu G, Li W, Gao X, Li X, Jurgensen C, Park HT, Shin NY, Yu J, He ML, Hanks SK, Wu JY, Guan KL and Rao Y (2007). p130(CAS) is required for netrin signaling and commissural axon guidance. J Neurosci 27:957-68.
3、Li X, Xue Gao X, Liu G, Xiong W-C, Wu J, and Rao Y (2008). Netrin signal transduction: roles of the guanine nucleotide exchange factor DOCK180 in attractive signaling. Nature Neurosci. 11:28-35.
4、Liu G, Li W, Wang L, Kar A, Guan KL, Rao Y, and Wu JY. (2009). DSCAM functions as a netrin receptor in commissural axon pathfinding. Proc Natl Acad Sci USA 106:2951-6.


0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号