阅读:0
听报道

视紫红质的发现
导 读
未来一周,2021年诺贝尔奖即将揭晓,刚刚获得拉斯克奖的光遗传学,再次成为诺奖热门之选。本文对光遗传学诞生过程中的一些重要发现进行了梳理。
撰文|郭晓强(石家庄职业技术学院副教授)
光可被细菌、藻类等低等生命和人类等高等动物通过视紫红质系统而感知。20世纪70年代后,几种细菌和藻类通道视紫红质的发现为光控系统的诞生奠定了基础。光遗传学最初由米森伯克于2002年首次实现并于2005年由迪塞罗斯(也译作代塞尔罗斯)和博伊登进一步完善,其应用极大地增强了对大脑功能的理解。
光遗传学可使科学家借助光来精确开闭特异神经元从而达到操纵神经元活性和动物行为的目的。光遗传学技术已被证明是在细胞和系统层面研究健康和病理大脑活性的一个非常强大且有用的工具。文章系统介绍了光遗传学诞生的历史背景、重大事件、发展过程、应用领域及重要价值等。
光对生命具有举足轻重的地位,“万物生长靠太阳”。对大部分植物而言,它们借助光合作用合成营养物质并释放出氧气,而动物则依靠这些营养物质和氧来维持生存。此外,光还可以指导细菌和植物的向光性,控制植物生长和开花时间。
对于人类和其他动物而言,借助光来观察和感知这个 “光明” 世界。该过程由 “眼睛” 完成,称为视觉。大部分视觉健康的人都可通过眼睛清晰地观察到这个世界,看到周围的花花草草和五光十色的世界。那么,我们是如何观察到这些事物的呢?文艺复兴后,人们对光的本质进行探索,从而对光的成像机制有了新认识,自然对视觉形成机制也产生浓厚兴趣。
视紫红质
视觉研究可追溯到18世纪。荷兰科学家列文虎克(Antonie Philips van Leeuwenhoek)借助显微镜观察眼视网膜结构,鉴定出视网膜色素上皮细胞(retinal pigment epithelium,RPE)、视杆细胞和视锥细胞等,并推测这些细胞与视觉形成相关。1851年,德国解剖学和生理学家缪勒(Heinrich Müller,1820—1864)首次报道视网膜视杆细胞显红色这一现象 [1]。遗憾的是,缪勒错误地认为红色由血液造成。尽管如此,缪勒仍被看作视觉生理研究的先驱。缪勒在视觉生物学领域作出诸多贡献,如首次描述视网膜神经胶质细胞,这类细胞也因此获名“缪勒细胞”。
博尔(Franz Boll,1849—1879)是一位德国生理学家,对视觉形成具有浓厚兴趣。1876年11月,博尔也观察到红色视杆细胞,并认定红色源于其含有一类特殊物质,纠正了缪勒早期的错误。博尔还发现视杆细胞的红色受光影响,光照可导致红色褪去,而在暗处又重新恢复,进一步说明红色物质与视觉形成相关。遗憾的是,博尔的早逝(年仅30岁)使研究没有进一步开展。
1877年1月,博尔的同胞、另一位德国著名生理学家屈内(Wilhelm Friedrich Kühne,1837—1900)进一步纠正博尔的不足,认定视网膜感光物质应为紫红色,并创造 “视紫红质(rhodopsin)” 一词。屈内还取得另一项重大发现,即胆酸可使视杆细胞内的视紫红质释放到溶液里,并基于这一原理首次从牛视网膜完成视紫红质的纯化 [2],屈内也因此成为视觉生理领域的奠基人之一(图1)。虽然已确定视紫红质参与视觉形成,但具体分子机制仍不清晰,直到20世纪30年代才有突破。

图1 视紫红质的发现
视黄醛循环
1931 年, 美国眼科专家尤德金(Arthur Yudkin,1892—1957)开始对视网膜成分进行分析,发现其含有一种维生素A样物质。其实,人们很早就知道维生素A缺乏可影响视觉形成,最常见的一种疾病叫夜盲症,但对维生素A如何参与视觉却知之甚少。
1932 年, 美国生理学家瓦尔德(George Wald, 1906—1997)来到德国瓦伯格(Otto Heinrich Warburg,1931年诺贝尔生理学或医学奖获得者)实验室开始全面研究视紫红质。瓦尔德首先借助光谱分析法证明青蛙、绵羊、牛等完整视网膜中存在维生素A,接着使用氯仿提纯视紫红质,化学显色反应表明所含物质与维生素A非常相似。
为进一步证实结论,瓦尔德加入瑞士著名科学家卡雷尔(Paul Karrer,1937年诺贝尔化学奖获得者)的实验室,而卡雷尔分离并确定了维生素A的结构。经过3个月研究,瓦尔德最终确定视紫红质中确实含有维生素A,从而表明视紫红质包含两部分:视蛋白(opsin)和维生素A [3]。随后,瓦尔德又加入德国海德堡迈耶霍夫(Otto Fritz Meyerhof,1922年诺贝尔生理学或医学奖获得者)实验室继续开展视觉形成研究。
一次偶然事件为研究带来重大契机!当时正逢假期,许多实验室人员都去度假,恰在此时运抵300只青蛙。实验室助理原本想丢弃,而瓦尔德则主动要求留下来用作实验材料。瓦尔德从青蛙视网膜提取到足够量的视紫红质,进一步分析后惊奇地发现所含的维生素A与卡雷尔所得维生素A尽管大部分性质相似,但仍有些许差异,因此将这种物质重新命名为视网膜色素(retinene)。瓦尔德还发现视网膜色素与维生素A之间可发生转变,并通过后来详细的结构分析确定了两者间的差异,因此视网膜色素更名为视黄醛,而维生素A则称为视黄醇 [4]。
20世纪50年代,瓦尔德和同事经过近20年探索,最终解析出视觉形成的 “视黄醛循环” 机制:静息状态下,视杆细胞内视蛋白与11-顺视黄醛结合形成视紫红质;光线照射可使11-顺视黄醛发生异构化转变为全反式视黄醛,从而与视蛋白分离,这个过程激活视蛋白,启动下游信号转导最终到达大脑视觉中心;全反式视黄醛可被运输到视网膜色素上皮细胞内经过几步化学反应重新生成11-顺视黄醛;11-顺视黄醛回到视杆细胞再次与视蛋白结合形成视紫红质,从而完成一次视觉感知过程(图2)。瓦尔德的发现很好地诠释了视黄醛参与视觉形成的机制,因此他分享了1967年诺贝尔生理学或医学奖。
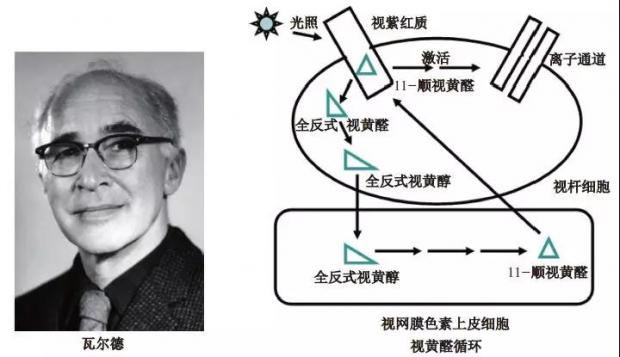
图2 瓦尔德与视黄醛循环
后续研究还揭示了视蛋白作用机制。视蛋白是一种G-蛋白偶联受体(G protein coupled receptor,GPCR)。光通过改变视黄醛结构而激活视蛋白后,可进一步使异三聚体G蛋白激活,从而使磷酸二脂酶活化,催化cGMP水解为5’-GMP而减少cGMP含量;细胞内受cGMP调控的离子通道关闭,导致细胞膜电位出现变化,最终传导至视觉中心而实现光的感知。
从这个过程可以看出,哺乳动物视紫红质的作用机制较为复杂,作为机体视觉感知过程尚可接受,如果将它们应用到其他系统则困难重重,因此有必要寻找更简单的感光系统 [5]。
细菌感光
最初认为只有高等动物才存在视觉系统,但这一观念在20世纪60年代发生改变。1967年,德裔美国生理学家斯托克尼乌斯(Walther Stoeckenius,1921—2013)成为加州大学旧金山分校的教授,重点研究生物膜(如红细胞膜和线粒体膜)结构 [5]。由于生物膜材料获取比较困难,具有电子显微镜背景的斯托克尼乌斯决定用生物化学方法研究盐生盐杆菌(Halobacterium halobium)细胞膜组成。随后两位新同事的到来壮大了实验室的力量。
厄斯特黑尔特(Dieter Oesterhelt,也译作奥斯特黑尔特)是一位训练有素的德国化学家,跟随吕南(Feodor Lynen,1964年诺贝尔生理学或医学奖获得者)获得博士学位,由于学术休假的缘故来到美国;布劳罗克(Allen Blaurock)是一位刚毕业的英国生物物理学家,原来在国王学院威尔金斯(Maurice Wilkin,1962年诺贝尔生理学或医学奖获得者)实验室从事X射线衍射研究 [6]。
厄斯特黑尔特和布劳罗克借助X射线衍射技术观测细菌细胞膜紫色组分时,意外观察到一种清晰的衍射图像,说明其含有一种高度有序的生物分子。厄斯特黑尔特还观察到紫色物质在添加有机溶剂后颜色变黄。此时,布劳罗克回忆起在国王学院研究青蛙视网膜过程中也观察到类似的颜色变化,这一提示促使厄斯特黑尔特大胆假设该物质可能也是视紫红质。为证实这一假说,首先需解答的问题是其含不含视黄醛。
从细菌中寻找视黄醛这一近乎疯狂的想法促使厄斯特黑尔特立即启动验证工作。借鉴青蛙视紫红质的研究方法,厄斯特黑尔特发现细菌的紫色物质具有类似的物理和化学性质,并且还含有视黄醛。基于这些特性,厄斯特黑尔特和斯托克尼乌斯于1971年确定这是一种新型视紫红质,根据来源将其命名为细菌视紫红质(bacteriorhodopsin,BR)(图3)[7]。

图3 细菌视紫红质
斯托克尼乌斯经过进一步研究后发现,细菌视紫红质是一种光依赖的离子通道。更大的突破在1975年,英国剑桥大学分子生物学实验室的亨德森(Richard Henderson,2017年诺贝尔化学奖获得者)解析了细菌视紫红质的三维结构,从而对视紫红质的作用有了更深入的认识。
1972年,重组DNA技术的发明为生命科学带来一场革命,同时也积极推动了细菌视紫红质研究的发展。研究人员将细菌视紫红质转入宿主细胞,结果发现光照可引起氢离子外流,从而证明其为一种光控的氢离子通道。1977年,研究人员在细菌中又发现另一种视紫红质——卤视紫红质(halorhodopsin),后续证明其介导氯离子细胞内流 [8]。
一系列的研究表明,即使简单如细菌这样的单细胞生物也存在 “视觉系统”,标志着一个新领域——低等生物视紫红质的诞生,从而促使科学家去寻找其他视紫红质。
藻类趋光
班贝格(Ernst Bamberg)是一位德国生物物理学家,从20世纪70年代开始研究细菌视紫红质的生物学功能,并利用体外实验证实BR是一种光激活氢离子通道。随着基因工程技术的发展和完善,生命科学的研究模式发生根本性改变,膜蛋白研究不再需要繁琐困难的提取过程,只需将外源基因在特定宿主细胞表达即可。
90年代,已加入德国法兰克福马普研究所的班贝格与从美国回来不久的德国电生理学家纳格尔(Georg Nagel)决定合作,共同研究细菌视紫红质在完整细胞中的生物功能。1995年,他们合作将细菌视紫红质基因成功转入非洲爪蟾卵母细胞,进一步精确证实光激活质子泵的电压依赖性 [9]。2001年,他们进一步在非洲爪蟾卵母细胞中证实卤视紫红质是一种氯离子通道(图4)。班贝格与纳格尔的合作一方面建立了视紫红质功能研究平台,另一方面也初显光遗传学雏形,即将外源视紫红质在靶细胞表达。

图4 藻类视紫红质
19世纪,绿藻(Chlamydomonas)等藻类就被发现具有向光性和受光调控的特性,但对这些现象背后的原因知之甚少。直到20世纪80年代,大量事实表明藻类也长 “眼睛”,即细胞膜存在感光物质,称为 “光受体”。
80年代初,德国生物物理学家赫格曼(Peter Hegemann)在博士就读期间就决定研究光受体。赫格曼和学生以莱茵衣藻(Chlamydomonas reinhardtii)为材料,借助电生理实验表明光的确可诱导藻类细胞产生电流 [10]。赫格曼决定采用生物化学方法将光受体蛋白纯化后研究其性质。遗憾的是,十余年辛苦努力最终以失败告终。根本原因在于光受体是一种膜蛋白,含量低、稳定性差且异质性高,这些都是蛋白质纯化的大忌。赫格曼不得不转换研究思路来解决这个难题。
2001年,绿藻基因组测序的完成为问题的解决带来转机。赫格曼通过全面搜索和比对绿藻基因组数据库,从中发现两个候选基因与细菌视紫红质具有较高同源性。
为加快研究进程,赫格曼决定寻求合作。他在获悉纳格尔的研究工作后,积极沟通并与其达成合作协议。赫格曼小组负责克隆两种绿藻视紫红质候选基因,并将其送给纳格尔开展功能研究;纳格尔则将基因转入人肾胚细胞HEK293并实现正确表达。功能研究表明,它们的活性均受光调控,并且介导阳离子如钠离子、钙离子等的摄入(图4),因此将其分别命名为通道视紫红质(channelrhodpsin,ChR)1和2 [11-12]。与ChR1相比,ChR2光激活时间更短,且离子通透性更强,因此更适合于研究。更为重要的是,赫格曼还推测这些通道视紫红质不仅可在普通细胞表达,而且也可在神经元中表达并影响电生理活性。这一论断直接催生了光遗传学。
至此,研究人员已经鉴定出三类光控视紫红质,分别是细菌视紫红质(介导氢离子输出)、通道视紫红质(介导阳离子输入)和卤视紫红质(介导氯离子输入)。它们在神经功能研究方面具有何种应用价值呢?这要从神经兴奋说起。
神经兴奋
大脑是神经系统的中枢,是机体最复杂和最神秘的器官。知觉、运动、兴奋、情感、语言、学习和记忆等过程基本都在大脑特异区域完成。大脑由上百亿神经元(亦称神经细胞)构成,这些神经元之间通过特定方式实现彼此间交流,以达到协调控制机体各种行为的目的。神经元活性受电信号影响。
正常情况下,神经元细胞膜内外两侧阴阳离子分布不均匀(这种现象称为极性):膜内钾离子浓度远高于膜外,膜外钠离子浓度又远高于膜内,最终形成一个外正内负的状态。未受刺激时(静息状态),规定膜外电位为0,则哺乳动物神经元膜内电位为负值,约-70mV,称为静息电位;外界刺激可导致离子通道打开,由于离子移动而引起膜两侧离子浓度发生变化,电位差也随之改变。如果-70mV向0方向改变,则称去极化(电位为0意味着内外无离子浓度差距,极化消失);相反,-70mV向更大负值变化则称超极化(意味着离子分布不均匀加剧)。
一般而言,去极化伴随神经元激活,而超极化则意味着神经元抑制,因此通过改变神经元细胞膜内外离子分布可实现精准控制神经元活性的目的。
1979年,美国索尔克研究所著名科学家、DNA双螺旋提出者之一克里克(Francis Crick,1962年诺贝尔生理学或医学奖获得者)在《科学美国人》发表一篇文章 [13],对脑科学未来的发展进行展望。古典神经生物学家通常采用电极刺激大脑特定区域神经元的方式来影响行为,克里克认为这种方法破坏性大且精确性不高,比如无法准确区分不同的神经元,这些因素导致所得结果准确性差。
为此,克里克提出应开发一种精确控制神经元活性的方法,允许研究根据需要只对特定神经元打开或关闭,同时不影响非相关神经元。具有分子生物学背景的克里克进一步指出可以对神经元细胞进行遗传改造,从而使它们可对外界信号(如光刺激)产生精准性应答。这一理念建立了光遗传学的思想雏形。
尽管光控细胞行为的理念已经提出,但真正实现则需要有可行的工具。2002年,这一想法终于首次变为现实。
神经光控
米森伯克(Gero Andreas Miesenböck)是一位奥地利神经科学家,跟随鲁斯曼(James Edward Rothman,2013年诺贝尔生理学或医学奖获得者)开展博士后研究。他主要借助荧光系统来检测神经元内囊泡运输,因而对光产生浓厚兴趣。
1999年,米森伯克建立自己的实验室,开始独立的科研生涯,目光锁定神经生物学。米森伯克对整个神经生物学领域一知半解,可以说有点 “门外汉” 的味道,但是恰恰这个因素反而使他在光遗传学方面首先完成突破,因为他不会受主流观点所羁绊。生命科学研究的基本策略在于首先控制某种因素(干预),然后依据结果确定因果关系,如敲除特定基因后动物出现某种表型异常(如个子变矮),据此可认为该基因参与了某个过程(如肢体发育)。
然而,由于神经系统自身的复杂性,长期以来神经生物学家主要依赖形态观察,而缺乏更多有效的干预手段。米森伯克想改变这一现状,他完全从一个生物学家的视点来看待这个问题,因此想为神经元安装一套感光系统(遗传学操作),然后借助光照(光学)来达到控制神经元的目的 [14]。为尽快实现这一目标,米森伯克邀请鲁斯曼的另一位学生、自己的师弟泽梅尔曼(Boris Valery Zemelman)加入团队,启动光控神经元活性的研究计划(图5)。

图5 米森伯克(左)和泽梅尔曼(右)
当时可用的感光系统主要是动物视紫红质,而哺乳动物过于复杂的光转导过程自然使米森伯克望而却步,不得不寻找其他替代品。幸运的是,他们在果蝇中找到一个简单系统,该系统只需三种蛋白参与,分别为视紫红质抑制蛋白-2(arrestin-2)、视紫红质(rhodopsin)和G蛋白a亚基(G protein alpha subunit)。他们为该系统杜撰一个名称 “chARGe”,之所以这样命名,原因在于 “charge” 还有另一层含义 “电荷”,说明该系统目的在于通过影响离子(带电荷的原子或原子团)运动而改变动作电位。
整个实验主要有两个关键步骤:首先将三种基因转入体外培养皿中生长的海马神经元;其次测试光照转基因神经元后动作电位的变化情况。2002年初,米森伯克小组首次在体外证明光照可控神经元的活性 [15],从而开启神经生物学研究的新篇章。
然而,果蝇chARGe系统在应用中存在诸多缺陷。首先需要三种蛋白,因此无论从转基因效率还是三者协调能力都将大打折扣;其次尽管可产生动作电位,但时间长、幅度小,不易观察和操作;最后,实验体系不稳定,产生的动作电位无规律,实验重复性差,从而也就意味着难以真正实现精确调控。
为解决这些不足,米森伯克小组对其进行改进。他们选择哺乳动物离子通道如P2X2和TRPV1等替代chARGe系统,这些通道蛋白都是单组分,因此大大降低实验难度,增加精准度。然而,这些通道蛋白活性本身并不受光调控,而是与配体如ATP和capsaicin等结合相关。米森伯克借鉴博士后工作期间的灵感,成功设计出光照影响二者结合的实验体系,从而达到光控神经元活性的目的 [16]。
但是,该系统也存在一些缺陷,如稳定性差(间接作用引起)等,还有一大不足在于这些离子通道都是哺乳动物蛋白,因此外源基因和自身基因之间存在互相干扰,这种现象称为系统不正交,从而为精准调控带来巨大难题。
2004年,美国加州大学伯克利分校克拉默(Richard Kramer)小组进一步对钾离子通道实现改造,最终也达到光控哺乳动物神经元活性的目的 [16]。然而,这些改进整体而言效果有限,因此迫切需要成分少、速度快、强度高、正交好的光控系统。藻类通道视紫红质恰好满足所有这些要求,因此,2002和2003年先后发表的藻类ChR文章立刻吸引了众多科学家的目光,激发他们快速加入该领域。
光遗传学
迪塞罗斯(Karl Alexander Deisseroth)是一位美国精神病学家和神经科学家,1998年获得斯坦福大学神经科学博士学位,主要研究神经元突触活动影响细胞基因表达的机制,而该研究对理解大脑的记忆功能具有十分重要的意义。
博士毕业后,迪塞罗斯最初想成为一名神经外科医生,因此在斯坦福医学院完成实习和精神科住院医师培训。然而,当他在精神分裂症患者治疗过程中体会到明显的无力感后,毅然决然改变选择,转向探索大脑的工作机制,以洞悉严重抑郁症、精神分裂症和孤独症等的发病过程,以期找到治疗良策。
随后,迪塞罗斯进入著名神经生物学家钱永佑(Richard Tsien)实验室继续博士后工作,并在这里结识了毕业于麻省理工学院拥有物理和电气工程背景的博伊登(Edward S. Boyden)。由于志趣相投,他们很快就成为科研挚友,并决定联合开发一种精准控制神经元行为的工具(图6)。他们最初设想采用磁控策略,未获成功。稍后博伊登将目光转向光控,对视紫红质产生浓厚兴趣,并获得细菌卤视紫红质质粒。
遗憾的是,因其他事宜这一计划未能实施。后来博伊登被米森伯克2002年发表的文章所深深触动,决定重拾光控神经元的研究计划。不久他又读到赫格曼发表的ChR2文章,很快就构思出一个完美的研究计划。

图6 迪塞罗斯(左)和博伊登(右)
2004年2月,博伊登建议迪塞罗斯从纳格尔那里借ChR2质粒。3月,迪塞罗斯与纳格尔达成合作协议,获得ChR2质粒使用权。质粒到达后,两人立即着手实验,但对实验能否成功仍然没有把握。
首先,ChR2在神经元能否高表达?其次,神经元是否也不需要视黄醛辅助?需要的话将无形增加系统复杂性。第三,外源ChR2能否产生足够电流而引发动作电位?最后,即使一切都顺利,会不会被其他实验室抢先发表?因为许多研究人员已经意识到光控的可行性。尽管有这些考量,但当务之急则是加快研究进程。
迪塞罗斯通过优化基因转染程序而使ChR2在海马神经元完美表达且正确定位于细胞膜,从而解除第一个担心;博伊登检测到表达外源ChR2的神经元电在不添加视黄醛的前提下,蓝光照射可在50 ms内引起足够阳离子流入,产生明显去极化效应。2015年1月,斯坦福大学本科生张峰(Feng Zhang)加入迪塞罗斯实验室,进一步构建出包含ChR2的病毒载体,大大增加了转染效率,电生理实验又进一步证实其高效的光控活性 [17]。
为取得发现优先权,他们于2005年4月19日迅速将论文提交到美国《科学》杂志,却被以 “没有在神经元中取得关于ChR2功能新发现” 的理由拒稿。5月5日,他们将文章转投英国《自然》杂志,也同样被拒稿。但幸运的是,稿件随后被转到《自然-神经科学》杂志,并最终被接收,于2005年8月在线发表 [18]。这篇文章也被看作光遗传学领域真正意义上的开山之作。
随后几个月,多家实验室先后发表文章证实光控神经元活性的可行性,包括日本的 Hiromo Yawo 实验室、美国凯斯西储大学的Lynn Landmesser和 Stefan Herlitze 的实验室,美国韦恩州立大学的潘卓华实验室(潘卓华的故事,详见“华裔光遗传学发明者不该被遗忘的故事”)。由于天然ChR2在实际应用中仍存在诸多不足,研究人员在此基础上对其进行改造。他们通过采取点突变的策略制备得到一系列新型ChR2变体,极大地丰富了光遗传学工具箱。2006年初,迪塞罗斯首次提出光遗传学一词 [19],用以描述联合光学和遗传学操作实现对特定神经元活性(或动物行为)精准控制的技术。
行为光控
体外光控神经元活性仅影响动作电位,而更大的挑战在于能否通过影响体内神经元动作电位进而控制生物行为。该领域的首个突破来自米森伯克实验室。已知果蝇存在逃跑反应,当它们感知周围存在威胁时会触发大脑特定神经元动作电位变化,进一步影响其他神经元活性并实施逃跑行为——飞走。
米森伯克的学生利马(Susana Lima)将光控蛋白基因转入特定神经元,随后的光照就会造成果蝇逃跑反应启动。然而,该实验体系存在一个明显的漏洞——果蝇本身对光线也非常敏感,单纯光照也可引起正常果蝇逃离。为区分果蝇的逃跑行为源于直接光控还是视觉感知引起的间接效应,米森伯克小组进一步将转基因果蝇头部去除以消除它们的视觉系统(果蝇仍可存活一定时间)。这些无头果蝇胸部仍保留神经元,光照后它们仍然呈现逃跑反应 [20],因而证实光确实直接控制神经元活性。
相对于低等动物果蝇,小鼠的精准行为光控就困难重重。首先,他们需要保证视蛋白只在特定神经元如参与睡眠、记忆或焦虑等表达。为此,他们分离得到神经元特异性启动子序列,将其连接到ChR2基因上游,从而保证ChR2只在这类神经元表达。其次,需要解决光照到达的问题。为此他们设计了一套设备,有效解决了这一难题。
2005年,迪塞罗斯小组开始尝试光控动物行为实验。他们首先成功制备下丘脑睡眠相关神经元特异启动子的ChR2转基因小鼠,然后将该小鼠放置于黑暗房间并处于睡眠状态,进一步蓝光刺激可唤醒小鼠,而正常小鼠则没有这种应答 [21]。随后,多个研究小组证实在动物水平上可实现光照控制行为的目的(图7)。

图7 光遗传学动物[14,25]
重大应用
光遗传学通过融合光学和遗传学的操作技术,实现对大脑特定神经元的精准时空控制。时间方面,控制精度可达毫秒量级;而空间方面,则可针对单一类型细胞。这一优势使光遗传学在探索大脑行为机制和部分疾病治疗方面具有得天独厚的优势。今天,全世界有上千家实验室在使用光遗传学技术研究大脑行为及其他科学问题。
在基础研究方面,神经生物学家借助光遗传学对许多大脑功能,如学习、记忆、新陈代谢、饥饿、睡眠、奖励、动机、恐惧、嗅觉和触觉等过程有了更为清晰的理解和认识 [22],并且揭示众多新机制,如个体特定状态下(如焦虑、风险规避等)大脑回路复杂行为的产生过程。
光遗传学技术还揭示帕金森病、阿尔茨海默病、癫痫等神经系统疾病发生的新机制,从而为药物研发提供重要靶点,同时也为新药筛选和测试提供重要技术平台 [23]。临床应用方面,光遗传学有望在失明等疾病治疗方面发挥重要作用。
未来几年,随着光遗传学技术的发展和完善,其价值将得到进一步体现,有望从根本上提升对神经系统以及非神经系统相关生理和病理过程的理解和认识,为部分疾病的治疗带来新突破。最近,光遗传学还在非人类灵长类动物中开展相关研究,从而给未来临床应用带来更大期望。当然,光遗传学本身涉及转基因过程,因此安全性和有效性问题尚待进一步评估。
光遗传学在几年内已经发展成为一种最先进的技术,并给神经生物学带来一场革命 [24]。2010年,光遗传学被《自然-方法学》评为 “年度方法”,还被《科学》杂志评为十年重大突破 [25-26]。这项技术突破使我们能够解决以前无法想象的生物学问题。
光遗传学被认为是自20世纪50年代以来神经科学领域最重大的进展之一,可与脑电极使用、功能性磁共振成像相媲美。光遗传学还直接推动2013年美国启动脑科学计划,大约耗资3亿美元,旨在开发治疗阿尔茨海默病、自闭症、精神分裂症和创伤性脑损伤等神经疾病的药物或技术。
光遗传学的成就已得到学术界广泛认可。许多作出卓越贡献的科学家,如厄斯特黑尔特、班贝格、纳格尔、赫格曼、米森伯克、泽梅尔曼、迪塞罗斯和博伊登等都先后获得多项科学大奖,包括欧洲脑科学奖(2013年)[24]、美国科学突破奖(2014年)和加拿大加德纳奖(2018年)[27]等。可以预见,部分科学家还有望分享诺贝尔奖。由于神经生物学一直是诺贝尔生理学或医学奖青睐的重点领域(10年内平均1~2项),光遗传学获诺贝尔奖也仅仅是时间问题。
科学启示
通过光遗传学的发展历程,我们还可以得到如下启示:
第一,微生物王国蕴藏着无尽宝藏。从限制性内切酶的发现及应用(1978年诺贝尔生理学或医学奖和1980年诺贝尔化学奖),到耐高温DNA聚合酶的使用(1993年诺贝尔化学奖),到视蛋白在光遗传学的价值,再到今天基因编辑技术——CRISPR-Cas9系统,无一不是将微生物成分借助基因工程应用到哺乳动物。毫不夸张地说,基因工程及相关技术的快速发展,微生物至少有一半以上的贡献。
第二,基础科学对技术革新意义重大。没有上百年来对光受体孜孜不倦的研究,没有20世纪70年代细菌视紫红质的发现和功能研究,没有藻类ChR的基因克隆,就不会有光遗传学的诞生,因此,强化基础研究对新技术的发明和完善具有十分重要的意义。
第三,技术进步对科学发展的推动作用。在知识日新月异的今天,许多科学问题的解决一方面依赖思维拓展,另一方面需要技术推进。尽管光遗传学的临床应用尚需时日,但这种技术手段对脑功能研究发挥了无法替代的作用,成为推动神经科学发展的重要工具。
本文原载于《自然杂志》2019年第3期,《知识分子》获权转载。
主要参考资料
[1] WOLF G. The discovery of the visual function of vitamin A [J]. J Nutr, 2001, 131(6): 1647-1650.
[2] COSTANZI S, SIEGEL J, TIKHONOVA IG, et al. Rhodopsin and the others: a historical perspective on structural studies of G proteincoupled receptors [J]. Curr Pharm Des, 2009, 15(35): 3994-4002.
[3] KRESGE N, SIMONI R D, HILL R L. Visual pigment molecules and retinol isomers: the work of George Wald [J]. J Biol Chem, 2005, 280(32): e29-e31.
[4] HOFMANN L, PALCZEWSKI K. The G protein-coupled receptor rhodopsin: a historical perspective [J]. Methods Mol Biol, 2015, 1271: 3-18.
[5] KRESGE N, SIMONI R D, HILL R L. Bacterial chimeras and reversible phosphorylation: the work of Walther Stoeckenius [J]. J Biol Chem, 2011, 286(31): e7-e8.
[6] GROTE M, O'MALLEY M A. Enlightening the life sciences: the history of halobacterial and microbial rhodopsin research [J]. FEMS Microbiol Rev, 2011, 35(6): 1082-1099.
[7] OESTERHELT D, STOECKENIUS W. Rhodopsin-like protein from the purple membrane of Halobacterium halobium [J]. Nat New Biol, 1971, 233(39): 149-152.
[8] HEGEMANN P, NAGEL G. From channelrhodopsins to optogenetics [J]. EMBO Mol Med, 2013, 5(2): 173-176.
[9] HARZ H, HEGEMANN P. Rhodopsin-regulated calcium currents in Chlamydomonas [J]. Nature, 1991, 351(6326): 489-491.
[10] NAGEL G, MÖCKEL B, BÜLDT G, et al. Functional expression of bacteriorhodopsin in oocytes allows direct measurement of oltage dependence of light induced H+ pumping [J]. FEBS Lett, 1995, 377(2): 263-266.
[11] NAGEL G, OLLIG D, FUHRMANN M, et al. Channelrhodopsin-1: a light-gated proton channel in green algae [J]. Science, 2002, 296(5577): 2395-2398.
[12] NAGEL G, SZELLAS T, HUHN W, et al. Channelrhodopsin-2, adirectly light-gated cation-selective membrane channel [J]. Proc Natl Acad Sci USA, 2003, 100(24): 13940-13945.
[13] CRICK F H. Thinking about the brain [J]. Sci Am, 1979, 241(3): 219-232.
[14] MIESENBÖCK G. Lighting up the brain [J]. Sci Am, 2008, 299(4): 52-59.
[15] ZEMELMAN B V, LEE G A, NG M, et al. Selective photostimulation of genetically chARGed neurons [J]. Neuron, 2002, 33(1): 15-22.
[16] MILLER G. Optogenetics. Shining new light on neural circuits [J]. Science, 2006, 314(5806): 1674-1676.
[17] BOYDEN E S. A history of optogenetics: the development of tools for controlling brain circuits with light [J]. F1000 Biol Rep, 2011, 3: 11.
[18] BOYDEN E S, ZHANG F, BAMBERG E, et al. Millisecond-timescale, genetically targeted optical control of neural activity [J]. Nat Neurosci, 2005, 8(9): 1263-1268.
[19] DEISSEROTH K, FENG G, MAJEWSKA A K, et al. Next-generation optical technologies for illuminating genetically targeted brain circuits [J]. J Neurosci, 2006, 26(41): 10380-10386.
[20] LIMA S Q, MIESENBÖCK G. Remote control of behavior through genetically targeted photostimulation of neurons [J]. Cell, 2005, 121(1): 141-152.
[21] ADAMANTIDIS A R, ZHANG F, ARAVANIS A M, et al. Neural substrates of awakening probed with optogenetic control of hypocretin neurons [J]. Nature, 2007, 450(7168): 420-424.
[22] KIM CK, ADHIKARI A, DEISSEROTH K. Integration of optogenetics with complementary methodologies in systems neuroscience [J]. Nat Rev Neurosci, 2017, 18(4): 222-235.
[23] SONG C, KNÖPFEL T. Optogenetics enlightens neuroscience drug discovery [J]. Nat Rev Drug Discov, 2016, 15(2): 97-109.
[24] REINER A, ISACOFF E Y. The Brain Prize 2013: the optogenetics revolution [J]. Trends Neurosci, 2013, 36(10): 557-560.
[25] WILLIAMS S C, DEISSEROTH K. Optogenetics [J]. Proc Natl Acad Sci USA, 2013, 110(41): 16287.
[26] STAFF N. Insights of the decade. Stepping away from the trees for a look at the forest. Introduction [J]. Science, 2010, 330(6011): 1612-1613.
[27] JOSSELYN S A. The past, present and future of light-gated ion channels and optogenetics [J]. eLife, 2018, 7: e42367.
[28] ISHIZUKA T, KAKUDA M, ARAKI R, et al. Kinetic evaluation of photosensitivity in genetically engineered neurons expressing green algae light-gated channels [J]. Neurosci Res, 2006, 54(2):85-94.
[29] LI X, GUTIERREZ DV, HANSON MG, et al. Fast noninvasive activation and inhibition of neural and network activity by vertebrate rhodopsin and green algae channelrhodopsin [J]. Proc Natl Acad Sci USA, 2005,102(49):17816-21.
[30] BI A, CUI J, MA YP, et al H. Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration [J]. Neuron, 2006, 50(1):23-33.
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}