- 编者按 -
在资讯发达的今天,或许你的手机上经常出现这样的推送:科学家发现抑郁症的分子机制、科学家发现调控睡眠或调控肥胖的基因……
“基因能调控行为”,这当然是毋庸置疑的,例如,period 基因调控人的生物钟。但当你看到上述新闻时,是否会纳闷:真的存在像电灯开关一样,能精准调控某些特定复杂行为的基因吗?比如,敲除或改变某个调控肥胖的基因,那无论每天吃再多,也不会长胖。
多年的研究使得科学家们发现,果蝇身上或许存在这样的 “开关基因”,这个基因能调控果蝇的求偶行为和性取向:果蝇撩不撩妹,撩妹还是 “撩弟”,都是它说了算。
撰文 | 潘玉峰
责编 | 王雨丹
调控行为的 “开关” 基因是真实存在的吗?
我们人体的任何行为都是由复杂的基因网络调控的,并不存在单个基因控制某个行为的情况。不过,可能存在某个基因,在行为调控的复杂基因网络中处于中心的地位。这种情况下,缺失该基因可能会导致相关行为的丧失(关),而在另一个没有该行为的系统中植入该基因,则可诱导该行为的表现(开),即 “开关基因”。
这样一个猜想在进化论的理论框架下是可行的,即在生命演化过程中,存在通过基因突变获得或者丧失某些行为的可能;同时,该猜想触发我们去研究更为简单的基因控制行为的模型(即使并不像 “开关” 那么简单),以提供更为基本的科学原理。
如果真的存在这样的 “开关基因”,那我们该如何把它们找出来呢?那些并非生存与繁衍绝对必要的高级认知行为,较大程度受到后天环境与经历的影响,因此受单基因的开关调控的概率较低。那么,与生存繁衍相关的本能行为,比如动物的求偶与交配、取食、睡眠、打斗、逃避天敌等,是否可能受到某些基因的开关调控呢?试想,某个单基因缺失,会导致取食或者睡眠的完全丧失吗?
生命演化应该不会选择这样的调控机制。但是,对于那些存在种属特异或者性别特异的本能行为,比如求偶与打斗行为,由于这些行为本身就具备性别或者种属差异,可能在进化过程中由特定的基因突变产生,从而受到特定基因的开关调控。
性别特异的基因与行为
两性的存在,提供了天然的具备性别差异的行为,特别是动物的求偶与交配行为。
自然界的动物很多都是雄性求偶(如熊猫、孔雀、狮子等),且它们求偶时通常会表现出一套求偶仪式,包括唱歌(发出特定的音频)、跳舞(特定的姿态)、展现美貌或者战斗力等。与此对应的是,雌性往往不能表现这些求偶仪式,而只是通过判断雄性的求偶质量来决定 “是否跟他谈恋爱”。
这种性别特异的求偶行为,由于其天然存在 “开与关” 的状态,因而具备受到特定 “开关” 基因调控的可能性。
在生命科学、人类医药与健康研究领域,一些动物生命活动中的生理和病理过程能在实验中与人或其他动物作为参照,它们被称为模式动物(如小鼠、斑马鱼、果蝇等)。我们可以通过一些模式动物来直观地感受基因调控的 “运作” 方式。
例如,在实验室常见的模式动物小鼠中,位于Y染色体上的Sry 基因决定了性腺的分化,由雄性睾丸分泌的睾酮或是雌性卵巢分泌的雌激素和孕酮,通过其受体调控神经系统的性别分化,进而调控行为的性别差异(图1A)[1]。
在另一种模式动物果蝇中,性别调控并非主要由Y染色体控制,而是由X染色体与常染色体的比值决定,再通过一套特定的分子调控机制,使位于X染色体的Sex lethal(Sxl)基因仅在雌性中表达蛋白SXL,而在雄性中则不表达,并最终调控了两个关键基因 fruitless(fru)和doublesex(dsx)性别特异的剪接方式,产生雄性特异的FRUM和DSXM蛋白和雌性特异的DSXF蛋白,并分别决定了雌雄的体细胞分化和性行为(图1B,1C)[2]。
因此,小鼠与果蝇的两性发育与行为分化,均由性别特异的基因表达方式调控;不同的是,果蝇的fru 和dsx 基因均以细胞自主的方式发挥调控作用,而在小鼠中则主要依靠性激素的远程作用。
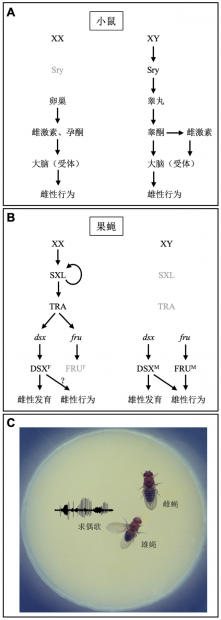
图1. 性别特异表达的基因与行为。(A)小鼠的性别决定通路。(B)果蝇的性别决定通路。(C)果蝇的求偶行为。其中仅雄蝇求偶,包括伸展单侧翅膀唱求偶歌;而雌蝇不求偶,其在性行为中的表现主要是接受或者拒绝雄蝇。
fruitless : 雄性果蝇求偶行为的潜在 “开关基因”
值得一提的是,果蝇求偶也是 “男性主动” 型,雄蝇的 “撩妹” 法包括追踪雌蝇、伸出前腿轻触雌蝇、伸展单侧翅膀唱求偶歌、伸喙舔舐雌蝇、弯曲腹部并尝试与雌蝇交配等固定步骤。雄性果蝇的求偶行为是本能行为,即它们不需要任何学习就能表现以上所有行为。
1963年,耶鲁大学的 Kulbir Gill 发现一类由X射线诱导的突变体果蝇不能与雌蝇交配,但却表现出很强的同性求偶行为,于是他将这一突变基因命名为 “fruity ”。由于 “fruity ” 一词在美国俚语中隐含对同性恋的歧视,同期也研究该突变的 Jeffery Hall(2017年诺贝尔生理学或医学奖获得者)便将其更名为 “fruitless ”,简写依然是fru,也代表了该突变体不育的表型,并沿用至今。
fru 被发现之后的很长一段时间,并未受到广泛重视。直到33年后的1996年,美国斯坦福大学的 Bruce Baker 实验室和日本东北大学的 aisuke Yamamoto 实验室几乎同一时期克隆了fruitless 基因,并发现该基因受到性别决定通路的调控。两性中相同的fru DNA产生了性别特异的fru mRNA,其中雄性特异的fru mRNA编码了目前已知的FRUM蛋白,而雌性特异的fru mRNA由于翻译过程中终止密码子未被剪切,因此未能产生有功能的FRU蛋白 [3, 4]。他们同时发现,FRUM蛋白特异地表达在神经系统中,且缺失该蛋白的果蝇均表现出不同程度的求偶行为缺陷。
2001年,Bruce Baker 等人在Cell 上发表综述,提出本文要讨论的核心问题,即复杂行为是否可能受到特定基因的控制,并提出fru 基因可能是一个基因控制行为的理想模型 [5]。在该论文中,Baker等人认为目前的研究已经显示出fru 对于雄性特异的求偶行为是必要的(图2),如果还能证明fru可以让另一个原本没有FRUM的系统(即雌蝇神经系统)也获得求偶行为的能力,也就证明了fru对于求偶行为是 “必要且充分” 的。
尽管他们并不认为这种单基因决定复杂行为的案例具备普遍性,但他们相信:单基因可能决定复杂行为的某个方面/模块;而像fru基因这样几乎能调控求偶行为所有方面的例子,将为研究基因如何调控复杂行为提供难以比拟的模型。
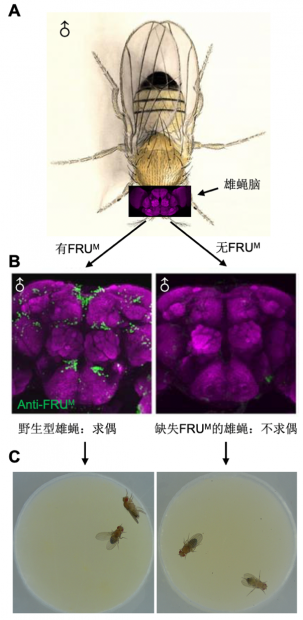
图2. fru对于本能求偶行为是必要的。(A)雄蝇及其脑结构。(B-C)野生型雄蝇脑内表达FRUM蛋白,并表现求偶行为;fru基因突变后不表达FRUM蛋白,雄蝇也丧失本能求偶行为。
2005年,Bruce Baker 实验室和 Barry Dickson 实验室分别在Nature 和Cell(2篇)上发表研究论文,通过同源重组的方法构建了fru相关的多种基因敲入果蝇,发现fru基因的缺失会导致求偶行为的丧失,失活fru 表达的约2000个神经元也使得雄蝇不再求偶。更为惊人的是,在雌蝇中表达FRUM蛋白,使得原本不求偶的雌蝇也开始表现出求偶行为(图3)!也就是说,他们在基因和神经环路两个层面证明了fru 确实像调控求偶行为的 “开关” 分子 [6-8]。
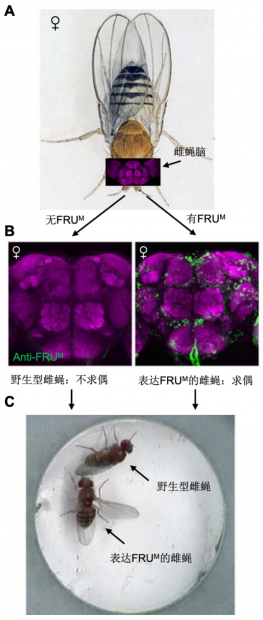
图3. fru 对于求偶行为是“充分”的。(A)雌蝇及其脑结构。(B-C)野生型雌蝇脑内不表达FRUM蛋白,不表现求偶行为;fru基因改造后在雌蝇脑内表达FRUM蛋白,雌蝇也开始表现求偶行为。
FRUM表达在约2000个神经元内,包含从感觉系统到中央整合、再到运动控制一整套调控求偶行为的神经环路。事实上,尽管研究fru 和求偶行为已经十余年,我仍然很难理解,为什么一个基因的表达和一种行为的神经环路几乎完全重合。不过,可以确信的是,fru 所标记的神经环路确实控制了求偶行为的几乎所有方面:失活所有fru 神经元,使得雄蝇完全不求偶;而人为激活fru 神经元,则使得雄蝇在没有雌蝇的情况下都能如上了发条般持续地表现求偶行为(视频1),直至死亡 [9]。因此,fru 神经元的活性与求偶行为的开与关也是对应的。
“同性恋基因”
在fru 基因的不同突变体中,有很多都能表现出同性恋行为(视频2),因此该基因也经常被报道为 “同性恋基因”。那么,fru 基因功能的改变或者缺失是如何导致同性求偶行为的呢?
由于来自同性或异性刺激的感受、整合均由fru 神经元调控,因此当fru 功能异常时,fru 所标记的求偶神经环路功能也会发生相应改变,而 “同性求偶” 也仅仅只是该神经环路功能异常表现之一。当然,并不是所有fru 相关的突变体都表现出同性求偶行为,比如某些fru 突变体雄蝇就是整体求偶能力下降。
值得一提的是,fru 调控求偶行为至少有两种机制。一是通过在发育过程中调控fru 神经元的发育,进而调控整个fru 神经环路的构建,使得雄蝇天生即能向雌蝇求偶;另一个机制是其在成年阶段发挥调控神经环路功能的生理作用,并抑制同性求偶行为 [10]。因此,fru 确实能够很大程度调控果蝇的性取向,但这也只是其调控求偶行为各个方面的一部分。
fruitless 并不是求偶行为的绝对开关分子
尽管fru 能控制果蝇的本能求偶行为,但其并不是严格意义上的 “开关分子”。2014年我们就发现,fru 对果蝇的求偶行为而言并非不可或缺,而仅是本能求偶行为所必要的:完全缺失fru的雄蝇,在单独饲养(没有后天经历)的条件下,确实不能求偶;但是在与其他果蝇群居几天之后,它们便能 “有样学样”,习得部分求偶行为,且这种后天习得求偶行为的潜力,受性别通路上另一个更为保守的dsx基因控制 [11]。
事实上,fru 所标记的约2000个神经元组成的求偶神经环路,在缺失fru 之后尽管会有一些发育上的缺陷,但是绝大部分神经元都还是存在的。这提示我们:果蝇体内应该存在着一个更为原始的、没有fru 但有dsx 的神经环路,该环路虽不具备本能求偶的能力,但是加上后天环境的作用,可能发生进一步的环路重塑,进而使果蝇拥有求偶行为。这对 “行为源自本能获得还是后天环境获得” 这一问题提供了深刻的启示。
虽然fru 不是求偶行为的绝对开关分子,但它对果蝇的求偶行为而言依然至关重要。万事有度,fru 并不能凭空塑造一套复杂行为,其功能本来就是建立在一套原始的求偶神经环路之上的;而这套原始的求偶神经环路可能受dsx 基因控制 [11]。换句话说,进化上更为保守的dsx 基因可能构建了一个尚不能工作的原始求偶环路,这套神经环路可以被后天环境所重塑并激活使用(后天习得求偶),也可以被fru 基因调控并在发育过程中构建完成(本能求偶)。
我们最近的研究还发现,fru 的表达强度或者表达位置不同,也能改变求偶行为的模式,产生天生异性恋、同性恋或者双性恋的果蝇(图4)。
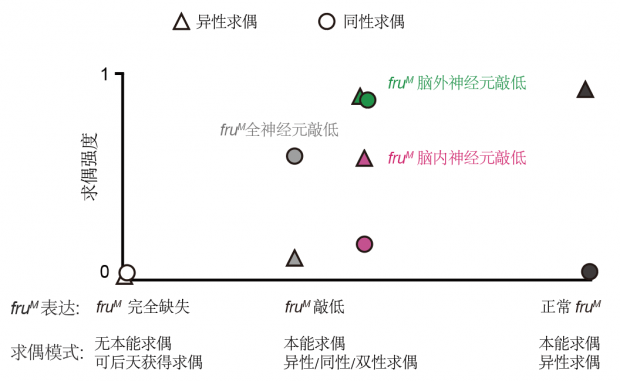
图4. fru决定了求偶行为的模式:本能异性求偶(正常fru)、后天习得性求偶(无fru)、本能同性求偶(fru整体表达低)、本能双性求偶(fru在部分神经元表达低)。
“开关基因” 研究的启示
从发现fruitless 距今已经快50年了。直到今天,还有很多实验室(包括我们)在研究fru 及其所标记的神经环路的功能。如此这般劳神费力,是否值得?作为一个并不能带来显著社会和经济效益的基础研究模型,它的启示,也许是让我们更好地认识自己,和我们所处的自然界。
首先,确实存在 “开关基因” 可以调控复杂行为,特别是某些本能行为(或者该行为的某一个层面),从而扩展了基因调控行为的深度;其次,fru 的性别差异表达及其对两性行为差异的调控,深化了我们对两性行为差异的机制性认识,这种认识将帮助我们更好地理解和尊重性别差异;再次,fru 与后天环境均能诱导求偶行为的产生,将帮助我们更加深刻地理解行为如何先天或是后天获得;最后,一个基因,通过其表达模式的改变,赋予同一个神经环路截然不同的功能(不同的求偶模式,图4),将深刻改变神经环路功能研究的内涵,即从研究功能转变为功能如何被调节,这对于理解进而调控行为输出至关重要,也是我们未来的主要研究方向。
作者简介
潘玉峰,东南大学生命科学与技术学院教授,研究方向为动物本能行为的神经调控。
”参考资料:(可上下滑动浏览)
1.Yang, C.F. and N.M. Shah, Representing sex in the brain, one module at a time. Neuron, 2014. 82(2): p. 261-278.
2.Yamamoto, D. and M. Koganezawa, Genes and circuits of courtship behaviour in Drosophila males. Nature Reviews Neuroscience, 2013. 14(10): p. 681-692.
3.Ito, H., K. Fujitani, K. Usui, K. Shimizu-Nishikawa, S. Tanaka, and D. Yamamoto, Sexual orientation in Drosophila is altered by the satori mutation in the sex-determination gene fruitless that encodes a zinc finger protein with a BTB domain. Proceedings of the National Academy of Sciences, 1996. 93(18): p. 9687-9692.
4.Ryner, L.C., S.F. Goodwin, D.H. Castrillon, A. Anand, A. Villella, B.S. Baker, J.C. Hall, B.J. Taylor, and S.A. Wasserman, Control of male sexual behavior and sexual orientation in Drosophila by the fruitless gene. Cell, 1996. 87(6): p. 1079-89.
5.Baker, B.S., B.J. Taylor, and J.C. Hall, Are Complex Behaviors Specified by Dedicated Regulatory Genes? Reasoning from Drosophila. Cell, 2001. 105(1): p. 13-24.
6.Demir, E. and B.J. Dickson, fruitless splicing specifies male courtship behavior in Drosophila. Cell, 2005. 121(5): p. 785-94.
7.Manoli, D.S., M. Foss, A. Villella, B.J. Taylor, J.C. Hall, and B.S. Baker, Male-specific fruitless specifies the neural substrates of Drosophila courtship behaviour. Nature, 2005. 436(7049): p. 395-400.
8.Stockinger, P., D. Kvitsiani, S. Rotkopf, L. Tirián, and B.J. Dickson, Neural Circuitry that Governs Drosophila Male Courtship Behavior. Cell, 2005. 121(5): p. 795-807.
9.Gao, C., C. Guo, Q. Peng, J. Cao, G. Shohat-Ophir, D. Liu, and Y. Pan, Sex and Death: Identification of Feedback Neuromodulation Balancing Reproduction and Survival. Neurosci Bull, 2020. 36(12): p. 1429-1440.
10.Chen, J., S. Jin, D. Chen, J. Cao, X. Ji, Q. Peng, and Y. Pan, fruitless tunes functional flexibility of courtship circuitry during development. Elife, 2021. 10.
11.Pan, Y. and B.S. Baker, Genetic identification and separation of innate and experience-dependent courtship behaviors in Drosophila. Cell, 2014. 156(1-2): p. 236-48.
制版编辑 | 卢卡斯

0
推荐
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}