阅读:0
听报道

作者说
今年的诺贝尔生理学或医学奖奖给了三位美国生物学家:遗传学家杰弗里·霍尔(Jeffrey C. Hall),分子生物学家迈克尔·罗斯巴什(Michael Rosbash)以及迈克尔·杨(Michael W. Young)。奖励他们在发现控制生物昼夜节律的分子机制上做出的重要贡献。
生物昼夜节律是众多生物节律的一种,也就是我们通常说的“生物钟”。昼夜节律(circadian rhythm)是一种以昼夜24小时为周期的生物钟,是动植物中普遍存在的基本的生命现象,揭示生物钟的分子构成和运行机理对我们了解生命、生命的演化和生命的活动具有重大的理论意义,同时也对治疗由于生物节律失常造成的人类疾病有着重要的应用价值。由于生物节律性活动是生物的一个基本行为,对生物钟行为的机理研究也首次建立了行为和基因之间的关系,克隆了第一个行为基因,为行为的遗传学和分子生物学研究奠定了基础。
我很有幸在1983年作为研究生在生物钟机理研究的初期加入了迈克尔·罗斯巴什的实验室,并和隔壁的杰弗里·霍尔实验室合作共同开展了果蝇生物钟基因的克隆和鉴定工作,参与并见证了这一历史事件的过程。我当时主要的贡献是在实验室里建立了果蝇的转基因技术,帮助鉴定了第一个生物钟基因: period(per)基因,克隆和鉴定了per基因的突变体和突变位点,并揭示了per基因可能参与了不同周期的生物钟的运行和调控。相关工作发表在PNAS和Nature等杂志上。当时在罗斯巴什实验室的另一个中国学生,我的好朋友刘欣也做出了重要的贡献。
撰文 | 俞强(中国科学院上海药物研究所研究员)
● ● ●
世界是物质的,物质是运动的,物质的周期性运动就是钟,生命物质的周期性运动就是生物钟。
什么是钟?什么是时间?
要了解生物钟,先要了解钟。什么是钟?这听起来是一个很简单的问题。我们都知道钟是一个人造的计时工具,我们每个人也都很熟悉钟。我们家里到处摆满了大大小小、各式各样的钟,我们的身上也佩戴着千姿百态、风情万种的“钟”。我们每天早上被钟叫起了床,被钟催着去吃饭,被钟赶着去上班。晚上盼望着钟敲响下班的铃声,等待着钟告诉我们什么时候该睡觉了。我们焦急地看着钟,期盼幸福的到来,我们无奈地数着钟走向生命的终点。我们制造钟、拥有钟,是钟的主人;我们被钟安排和指挥着每天的生活,又是钟的奴隶。我们在钟声中诞生,也在钟声中消失,我们的整个生命都在钟的掌控之中。
钟到底是什么?它,为什么有那么神奇的力量?我们为什么需要钟?我们的生活为什么要让钟来安排?我们的生命为什么要被它掌控?当我们仔细地深度思考这些关于钟的问题时,想给它一个精确的、科学的定义和哲学的解释时,我们会发觉我们对钟了解得太少了。我们除了知道人造的钟是一个计时的工具之外,对自然的钟,对钟的自然本质和本原意义,对钟在我们生命中的功能和对这个世界的意义,我们知道和了解得太少了。
钟,究竟是什么呢?什么是自然的钟?什么是生物的钟?人造的钟和自然的钟又是什么关系?让我们揭开钟的神秘面纱,走进钟的内芯,去探索一下钟的本质。
我们生活的这个世界是一个物质的世界,是一个在空间和时间中充满了我们看得见和看不见、测得到和测不到、大小不同和形态各异的物质的世界。这些物质大到无边无际的宇宙,小到无限可分的原子粒子,在形态上有似乎“亘古不变”的“死物”,也有生生不息、代代相传的生物。然而不管什么物质,都在空间和时间中不停地、无休止地运动着。
物质多种多样,物质的运动形式也千变万化,其中我们最熟悉的就是物质重复性的、以一定时间为周期的、有节律的运动。我们能测到的最大的物质,如宇宙中的每个星系,都在各自轨道上有节奏地运行;我们能测到的最小的物质,如基本粒子,它也是构成世界上所有物质的基本单元,它的自旋和衰变所产生的辐射波都是律动。如果说物质的单元是基本粒子,那可以说物质运动形式的单元就是律动。这种物质周期性律动的现象、机制和功能就是“钟”。钟,既是物质,也是物质的属性和运动形式。“钟”性,即韵律、节奏,是物质的普遍属性,可以说世界上每个物质都在时间上作不同形式的、周期性的运动。这些律动的周期可以无限小,以秒、纳秒、渺秒记,也可以无限大,以年、千年、亿年记。我们人造的钟不过是人类按照自然规律,模仿自然而制造的反映物质特性和自然规律的一个测量时间的装置,是自然规律的镜子。
如果说人造的钟是模仿自然、测量自然时间的装置,那时间又是什么呢?我们该如何给时间下个定义?
时间和空间一样,是个很难定义的概念,也是让哲学家们伤脑筋的问题。时间不是一个物质,也不是一个让物质在其中运动的容器。公元 5世纪,古罗马帝国的思想家、神学家和哲学家圣 · 奥古斯丁(St Augustine)在谈到时间时说:“如果没有人问我,我知道什么是时间;如果有人问我什么是时间,我只能回答不知道。”理论物理学家、美国哥伦比亚大学教授布莱恩 · 葛林(Brian Greene)说:“给时间下定义的难处在于我们很难,甚至不可能不用时间这两个字来给时间下定义。”牛顿曾给时间下了一个定义(牛顿时间):时间是物质世界的一部分,是一个事件在其中依序发生但独立于事件的维度和量度。而爱因斯坦则更喜欢“时间就是钟所测量的那个东西”(Time is whatever clocks measure)。
综合起来我们可以试着给时间下一个定义:时间是我们生活其间的四维空间中的一维,是一个与三维立体空间并存的另一个坐标轴。时间是一种度,是可以用钟来测量的度;时间也是一种力,是一种阻止事件同时发生并保持事件有序发生的力。而在实际生活中,物质重复性的运动和事件的间隔被用来定义时间与时间的单位。这个重复性的运动和事件的律动,就是韵律和节奏。有了这个可操作的时间的定义,我们就可以用这个定义来制造测量时间的钟:一个自主的、做节奏性运动的物质或装置。
人造的钟作为计时器,从最早的日晷、滴漏、沙漏、机械摆钟,到现在的石英钟和原子钟,不论其形式和构造发生了多少变化,其原理都和自然的钟一样:律动;其核心都是一个:振荡器。目前世界上人类计量时间最精确的标准钟——原子钟,就是利用自然界的原子吸收或释放能量时发出的周期性辐射波频率来计时的。 1967年和1997年召开的国际度量衡大会把在绝对零度和零磁场环境下,处在基态的铯133原子在两个超精细能阶间跃迁所产生的辐射波经历9 192 631 770个振动周期的时间定为国际单位制(SI)中时间的基本单位——秒。这个人类计时的标准钟“铯钟”就是一个自然的钟,是自然物质的本来属性,是自然的规律。现在人们所制造和使用的钟都是以这个自然钟的韵律为标准来校时的。
因此,钟是一种物质,既是人造的物质,更是自然的物质。
钟是一种物质的运动形式,是大自然的节奏规律,是物质的普遍内在属性。
生物钟和生物钟的起源
生物,也是一种物质,只不过是一种“生”物,是能一物生一物的有“生”命的物质。生物钟,因此也就是万千自然物质钟的一种,也叫生物节律或生物韵律,是指不同生物体内各种随时间变化而做周期性变化的生理生化活动。广义的生物钟是指生物体所表现的所有的生物节律,如我们心脏的跳动、肺的呼吸,昆虫翅膀的扇动等。而平常所说的生物钟是指地球上的生命随地球的周期性运动而产生的各种周期性变化的生理生化活动。例如,以 24小时为周期的人的血压、体温、体力、情绪等生理指标的律动,植物花的开闭和叶片的光合作用等,都是随地球的自转而产生的昼夜变化律动。而众多生物以年为周期的、在春秋季的繁衍以及在冬季的冬眠则是由于光照时间随地球围绕太阳的公转而呈周期为一年的变化。绕地球旋转的月球又给地球上的环境,特别是海水的环境,造成了一个以月和半月等为周期的韵律,从而使海洋沿岸的动物产卵、排精等都具有相应的周期性。这种地球上的生命随地球和其他星球的节律性运动而表现出的节律性现象就是我们通常所说的生物钟。生物钟和非生物钟在运动形式上都是物质的周期性运动,但在机理上有着质的区别。非生物钟是物理力作用的结果,而生物钟则是生物化学反应的结果。
从生物的演化和自然选择原理来说,地球上所有的生命现象都是大自然对生物适应环境的演化选择的结果。由于生命约出现于地球形成 10亿年之后,从它诞生开始,生命最简单的生物分子之间的生物化学反应就受到地球上早已存在的各种物质和环境因素的影响,其后的变化也都在地球环境的掌控之中,因此地球上生命的一切活动都受到地球周期性律动的影响,所有的生物节律也都是环境对生物演化的选择的结果。从这个角度看生命,生物的节律性运动和生物钟应该是普遍的生命现象。
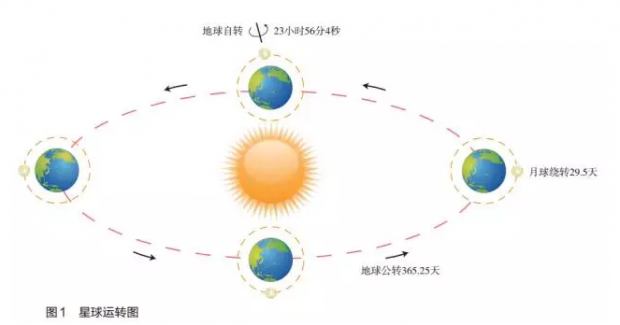
地球在宇宙中最明显的律动就是围绕太阳的公转和围绕自己轴心的自转。地球自转的周期为 23时 56分4秒,围太阳公转的周期为365.25天(图 1)。地球还有一个围着它绕转的卫星:月球。月球公转的周期为 29.5天(朔望月)。这样月球绕着地球转,地球和月球一起绕着太阳转,这些地球和地球环境的旋转律动无时无刻不在影响和控制着地球上包括生物在内的一切物质和物质的运动。而太阳是地球上所有生命的能源和动力,生命的运动不能不顺从于太阳能源的周期性律动而随之律动。这就让我们不难理解生物为什么会有生物钟,为什么生物钟的律动频率与地球及月球的律动频率是同步的。只有当生命的节奏与自然环境的节奏吻合的时候,生命才能够更好地生存,才能在生物演化的过程中被自然选择而保存下来。而以 24小时为周期的地球围绕太阳的律动是影响生命节奏的最重要的律动,因此地球上的生命活动也都跟随着能量来源的节奏昼夜 24小时不停地、有序地变化。这个生物周期为 24小时的节奏性运动称为日节律或昼夜节律(circadian rhythm)。
最常见的日节律就是植物的光合作用和动物的昼夜活动。植物的光合作用把太阳光的能量转化成了化学能,植物的化学能又被转化为动物的化学能。生物的食物链实际上就是一个能量的传递链,这条链的源头所吸收的是太阳光的能量。太阳光在地球上的律动导致了地球上食物链各个环节的律动,因此最普遍和常见的生物钟就是以日为周期的生物钟。由于太阳光一方面给生物提供能量,另一方面也对生物的大分子如 DNA有损伤作用,因此生物也演化出了为保护 DNA而躲避强光的生物钟行为,如动物为避免中午太阳的高温而在早晨和傍晚觅食的生物钟行为。生物除了昼夜节律,还有周期比日长的节律,叫做长日节律(infradian rhythm),如女性生理性活动的月节律,植物的四季变化,动物的生殖、迁徙和冬眠等年节律。动物还有周期比一日短的节律——短日节律(ultradian rhythm),如人的心脏跳动节律和呼吸节律。这些各种形式的生物的自主的节奏性变化和运动就是生物钟。
生物钟是自然对生命适应生存环境的一种生命的运动形式的选择,它是生存环境的律动在生命中的反映,因此生命的每种律动都有与之相对应的环境的律动。日节律、月节律、年节律是地球和月球在星际中转动的节律,也是地球上生物生命活动的最常见和最基本的节律,是生物最基本的钟。它告诉和指挥我们活动的时间,也告诉和安排我们休息的时间。
生物钟的发现
人类最早认识到的生物钟现象是人自身的周期性变化,以及植物的叶和花的朝夕节奏性变化。东方世界中国的古代哲学家对人体和自然的相互关系特别重视,早在公元前 300年的战国时期,《黄帝内经》就详细描述了人的身体状况随季节变动而变化的周期性现象以及人体的脉象节奏。植物是地球上早期出现的生命形式,也是食物链最底层的物种,它们的养料来源于阳光、空气和水,它们都需要太阳光能来进行光合作用,因此植物的昼夜节律运动是最普遍也是最明显的现象。古代有很多关于植物节奏性活动的记载。西方世界有记载的最早关于生物钟的故事也发生在公元前300多年,古马其顿国王亚历山大大帝手下的船长安德罗斯申尼斯(Androsthenes),曾向亚历山大大帝描述罗望子树的树叶昼夜开合的运动的现象。而正式提出和记载植物的生物钟行为的人是大家熟知的、以创建动植物分类和命名法而享誉世界的瑞典博物学家林奈(Carolus Linnaeus)。林奈在观察和研究植物时注意到菊花每天花开花闭的时间几乎一致,同时他注意到不同的花每天开闭的时间不一样。于是林奈就把不同的花在地上摆了一圈,不同的花在不同的时间花开花闭就构成了一个生物的报时钟。
全世界首先用科学的方法来研究生物节奏的是 18世纪的法国天文学家让—雅克 · 道托思 · 麦兰(Jean—Jacquesd’ Ortous de Mairan)。他在研究地球运转的同时注意到含羞草在白天时叶是张开的,但到了晚上就闭合了。于是他好奇地把含羞草放到一个不透光的盒子里,然后观察叶的变化,结果发现含羞草的叶在恒定的黑暗中仍然保持了它以 24小时为周期的昼夜变化。麦兰第一次记录了内源性的、而不是光或其他外因造成的昼夜节律性的振荡,无意中成为第一个发现日节律的人。麦兰的发现比日节律这个词的正规使用早了230年,并且他也是第一个用恒定环境来检测和判断一种生物节律是受外部刺激的还是内源性的人。
继麦兰之后,法国农学家亨利—路易斯 · 杜哈麦 · 芒修(Henri—Louis Duhamel du Monceau)做了一个类似的实验。他把含羞草放到了一个黑暗并保持温度恒定的环境中,发现含羞草的叶的昼夜变化仍然存在,从而发现了生物钟节律不仅不依赖于光,而且不依赖于温度,进一步提示内源性生物钟的存在。 1832年,瑞士植物学家艾尔芬斯 · 坎道拉(Alphonse de Candolle)发现,植物的叶在恒定的条件下的律动周期并不完全与地球的自转周期相同,有的植物律动周期比 24小时长一点,而另一些则短一点,这从另一方面提示生物钟的节律是独立于环境的、自主的节律,否则它们的节律周期应该和地球自转一样。
上述三人的工作都从不同的方面指向了一个自主的、内源性的生物钟的存在,但他们三人的工作在当时并没有引起其他科学家的注意。在他们之后的达尔文也做过类似的研究,并和他的儿子一起共同发表了他们对生物钟研究的结果。
近代对生物钟的广泛和深入的研究是从 20世纪中期开始的。德国生物学家欧文 · 本林(Erwin Bunning)、尤金 · 沃尔特 · 路德维格 · 阿绍夫(Jürgen Walther Ludwig Aschoff)和美国生物学家科林 · 皮登觉(Colin Pittendrigh)被认为是生物钟(也称时间生物学)研究领域的创始人。本林研究的是植物的叶闭合活动的生物钟行为;阿绍夫研究的是人的体温、活动等和鸟类的一些生物钟行为;而皮登觉研究的则是果蝇运动的生物钟行为。三个人所研究的生物系统虽然不同,但三个人都总结出相同的关于生物钟的几个基本特征。
◇ 生物钟是内源的、自主的、不依赖于环境变化的生物节律。
◇ 昼夜节律的生物钟周期不是精确的24小时,而是接近于24小时。
◇ 生物钟具有温度补偿的性能,能在不同的温度条件下保持稳定。
◇ 光照不是产生节律的原因,但能够调节和重置昼夜节律生物钟的相并使其同步。
这个时期的生物钟研究虽然对生物钟存在的普遍性以及生物钟的某些基本特征和规律有了较为深入和广泛的了解,也知道怎样用光来重置生物钟的启动和用化合物来停止生物钟的运行,但此时的研究还基本停留在对现象的描述和对机理的猜想阶段,而对生物钟的元件和运行机理却还一无所知。生物学家们还无法从细胞和分子的水平来想象一个生物钟的构造和生物化学反应的原理和过程。
生物钟在生物体中的部位
在明确生物钟的现象后,科学家自然想知道生物钟是什么样的?它在生物体内的什么部位?由哪些组织和细胞构成?这个钟是如何工作的?生物体内有一个钟还是多个钟?不同的钟之间是如何配合工作的?
对于生物现象的许多机理研究都是在大鼠、小鼠和果蝇这三种动物模型中展开的,它们也同样被用来研究生物钟的机理。美国约翰 · 霍普金斯大学的瑞科特(Curt Paul Richter)教授在 20世纪 60年代对大鼠的生物钟行为做了多方面的详细研究,在代谢、内分泌、神经等多个系统中探索了影响生物钟行为的元件和机理。他用手术的方法分别去掉了大鼠的肾上腺、性腺、脑垂体、甲状腺、松果体、胰腺等所有他能想到的腺体,也用了电击、振动、麻醉等各种处理,都没有能消除掉大鼠的生物节律。直到他在大鼠的大脑各个部位做了 200多次手术实验后,最终发现了大鼠下丘脑的前端是大鼠生物钟的中心。当他用手术的方法损伤了下丘脑前端的时候,他发现大鼠的多种生物节律被破坏了,因此瑞科特教授是世界上第一个指出下丘脑的前端可能是哺乳动物生物钟的振荡器所在的人。后来美国加州大学伯克利分校的朱可(Irving Zucker)教授和他的学生史泰芬(Friedrich Stephan)以及芝加哥大学的莫尔(Robert Moore)教授对下丘脑做了进一步的精确损伤研究,发现了下丘脑前端的视交叉上核是启动大鼠生物钟的关键元件。当他们人为地损伤了视交叉上核时,大鼠的内分泌节律和行为节律就丧失了,由此判定视交叉上核可能是大鼠生物钟的起搏器。
最终对视交叉上核的生物钟身份的确定是通过两个关键的体外和体内实验。日本东京大学(TheUniversity of Tokyo)的井上进一(Shin—Ichi Inouye)和川村宏(Hiroshi Kawamura)直接测量了视交叉上核神经细胞在体内和体外的电生理活动,发现视交叉上核神经细胞的电生理活动是以 24小时为周期的日节律活动,由此确定了视交叉上核为哺乳动物生物钟的振荡器。后来的许多实验进一步证明,哺乳动物的很多节律性行为和生理活动,如睡眠、运动、警觉、激素水平、体温、免疫功能、消化功能等,都受视交叉上核调控。如果没有视交叉上核,这些生物节律就都消失了。虽然后来的研究发现体内其他许多细胞和组织也都有它们自己的以 24小时为周期的生物钟,但视交叉上核起到了一个调控和协调周围组织的生物钟保持同步运行的作用,从而被称为“主钟”。
视交叉上核的结构和它在脑内的位置决定了它作为生物钟的起搏器和振荡器的合理性。一方面它是大脑中许多直接从视网膜接受神经信号的核之一,通过视网膜下丘脑束从视网膜上的一些光敏神经节细胞中接受信号;另一方面它和大脑的其他许多部分相互作用,将信号传递给大脑的其他部位。
视交叉上核由约 20 000个神经细胞所组成,位于视神经交叉的上面。它的背部是一群具有在无光的环境里也能保持自主的约 24小时节律活性的神经细胞;而它的腹部两侧则是两群活性受光调控的神经细胞。视网膜上含有黑视蛋白的神经节细胞通过视网膜下丘脑束直接和视交叉上核的腹部两侧相连接。有光照时,视交叉上核的腹部两侧神经细胞就会把光照的信号传递给整个视交叉上核,视交叉上核整合来自各方的信号,从而进行节律同步调节。一种有γ—氨基丁酸介入的机制被认为是起到了偶联视交叉上核的腹部区和背部区的神经细胞的作用。视交叉上核再把这些节奏性的信号传递到其他下丘脑的核群和松果体去调节体温以及产生皮质醇和褪黑素等激素。视交叉上核同时也产生多种神经递质和多肽,如血管加压素和血管活性肠肽。
生物钟是一个生物适应环境的古老的机制,从原理上讲,地球上所有的生物都生活在同一个周期为24小时的日光变化的环境中,并且每种生物都是从单细胞演化而来,因此每个生物体的每个细胞都可能有一个生物钟。事实似乎也证明正是如此。果蝇和鼠的生物钟被发现存在于多种组织中,小鼠在体外生长了 30年的成纤维细胞株中的生物钟基因的表达仍然呈现周期为 24小时的节律性变化。从古老的单细胞生物如蓝藻,到复杂动物如人,在各种不同生物的不同细胞中,都有生物钟的存在。不同细胞中的生物钟各自分管着不同的节律性功能,而复杂动物大脑中的生物钟,起到了协同整合和介导光的调控的作用。
在人等哺乳动物中,下丘脑的视交叉上核就是这样一个协调周围组织“子钟”的“主钟”。大脑是人体的司令部,大脑中的生物钟也是人体的司令钟。
生物钟的分子机理
对生物钟的分子机理研究最早是在果蝇中展开的,动物中最早被阐明机理的生物振荡器是果蝇的日节律的生物振荡器,最早研究果蝇生物钟行为的科学家是美国加州理工学院的生物学家本哲(Seymour Benzer)教授。他和他的学生克洛普克(Ronald Konopka)在 20世纪70年代首先运用遗传学的方法,用化合物诱导了果蝇 DNA的突变,然后从发生了基因突变的子代果蝇中筛选到了生物钟行为发生了改变的果蝇突变种。这些变种果蝇有的是完全失去了日节律的变种,它们的活动在恒定无光的环境里变得毫无规律,叫“无周期突变”;有的虽然仍然保持规律性的运动,但它们的日节律周期变得长短不一,有的比24小时长,叫“长周期突变”;有的比 24小时短,叫“短周期突变”。而且,这些突变的性状是能够遗传的。这些遗传学实验揭示了生物钟基因的存在,首次向科学界证明果蝇的生物钟行为是基因调控的。本哲和克洛普克将他们发现的生物钟基因命名为“周期基因”(period/PER),并在果蝇的基因染色体上确定了PER基因的位置。这是世界上第一个被发现的生物钟基因。
最终证明和明确基因是生物钟行为的决定因素是生物钟基因的克隆。在美国布兰达斯大学(Brandeis University)的分子生物学教授罗斯巴什(Michael Rosbash)和遗传学教授霍耳(Jeff Hall)是一对篮球场上的好战友,球场让他俩经常有机会在一起讨论一些科学上的新发现。当本哲和克洛普克发表了他们关于果蝇生物钟突变种的时候,罗斯巴什和霍耳注意到了这一发现的重大意义,于是他们俩就决定合作来克隆这个果蝇的生物钟基因。经过了2年多的努力,罗斯巴什和霍耳的实验室终于在 1984年从果蝇中克隆到了第一个生物钟基因: period/PER基因。继 PER基因之后,另外几个生物钟元件基因 TIM、 CLK、CYC、VRI和PDP1等也相继被克隆。10年后,当时在美国西北大学(Northwestern University)的约瑟夫·高桥(Joseph Takahashi)教授又在1994年发现了小鼠的生物钟基因 CLK(CLOCK),并在 1997年克隆了 CLK基因,他成为发现哺乳动物生物钟基因的第一人。从此,生物钟的研究发生了根本的改变,从对现象的描述和对机理的猜测,进入在细胞和分子的水平上解析生物钟的元件和阐明机理的研究。
经过 30年的研究,科学家现在对动物中以 24小时为周期的生物钟的构成和机理已经有了基本了解。动物生物钟的循环基本上是一个基因表达的负反馈环路,是一个基因表达的振荡器和过程。在这个过程中有两个调控基因转录的异二聚体蛋白起了关键作用:一个是直接作用于 DNA促进转录的转录因子 CLK和 CYC的二聚体 CLK—CYC,另一个是抑制CLK—CYC转录功能的PER和 TIM的二聚体PER—TIM。 CLK—CYC的功能是促进一系列包括 PER—TIM在内的和生物钟行为相关的基因的表达。这些基因的启动子部位都有一段称为 E盒元件的DNA序列,CLK—CYC作用于 E盒序列促进这些基因的表达。表达后的 PER和 TIM蛋白先在细胞质中逐渐累积,到了晚上当两种蛋白累积达到一定的量后又被转运到细胞核中转而抑制 CLK—CYC的转录活性,从而抑制它们自己以及所有 CLK—CYC下游基因的表达,减少被表达的量。而在细胞质中的 PER蛋白被逐渐水解,从而构成了一个以 24小时为周期的负反馈基因转录和翻译的振荡(图 2)。
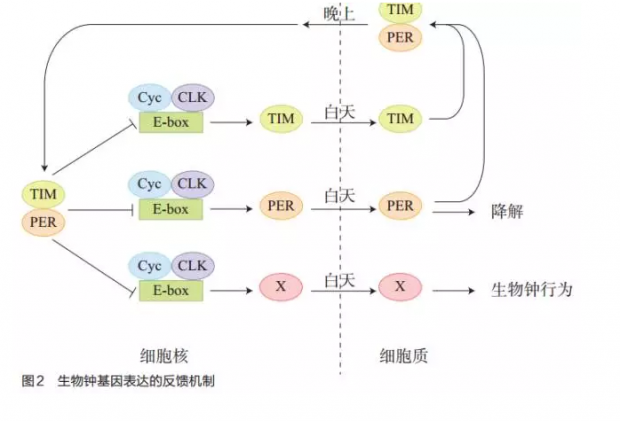
这种从 24小时为周期的节律具有一种特性,就是它的起始点可以被光照重新设置。这个重设置过程也是一个由蛋白质介导的生物化学过程。在果蝇中,这个有重设置功能的蛋白称为 cryptochrome(CRY)。CRY蛋白有感光的功能,它和 TIM的相互作用是光依赖的,并且这种相互作用的结果是 TIM的降解。而失去 TIM的 PER蛋白不稳定,最终也在有光照的白天被降解,其结果就是减少了对 CLK—CYC二聚体功能的抑制,从而使得 CLK—CYC介导的基因转录重新开始。
生物钟是普遍存在的,提示它在生物演化史中是一个古老的现象。果蝇和哺乳动物中的生物钟基因相似,但和植物及单细胞生物的基因不同。不同生物生物钟基因的不相似性提示生命的起源是多元平行的起源。虽然不同种生物的生物钟基因多种多样,但它们的工作原理都是类似的:基因表达的负反馈调节。这个生物振荡器就是所有生命所共有的、最基本的生物化学反应的振荡器:基因表达的振荡器。
生物钟的功能
所有的生物性状都是自然对生物适应环境的变化选择的结果。有利于生存和繁殖的性状就在生物演化的过程中被自然选择保留了下来,反之则被淘汰。生物钟也是一样。生物钟是生物在长年的演化过程中被环境选择出来的一种预见和预警机制,它预见一个规律性事件的发生,通常是食物和危险的出现。生物钟让一个生物个体预见到食物的定时出现而提前准备并及时到场;生物钟也预见不利于生理活动的事件,比如高温和寒冷的定时出现而提前规避。能掌握环境变化规律并预见环境变化的物种显然有生存和繁殖的优势,因此被自然所选择。生物钟的元件和机理就这样在长期的生物演化过程中被自然选择保留了下来,成为了普遍的生物现象。光照的亮度变化选择和保留了日节律生物钟的构造和机制,而光照长度的变化则选择和保留了年节律生物钟的构造和机制。
人体的生物钟以及和健康的关系
人的生物钟就是人体内随时间作周期变化的生理生化过程、形态结构以及行为等现象。人体内的生物钟多种多样,人体的各种生理指标,如脉搏、体温、血压、体力、情绪、智力等,都会随着昼夜变化做周期性变化。例如,体温早上4时最低,下午6时最高,相差有1 ℃多。
科学家经过多年的研究,已经对人体的许多生理生化活动的昼夜节律现象有了比较清楚的了解,比如(图3):
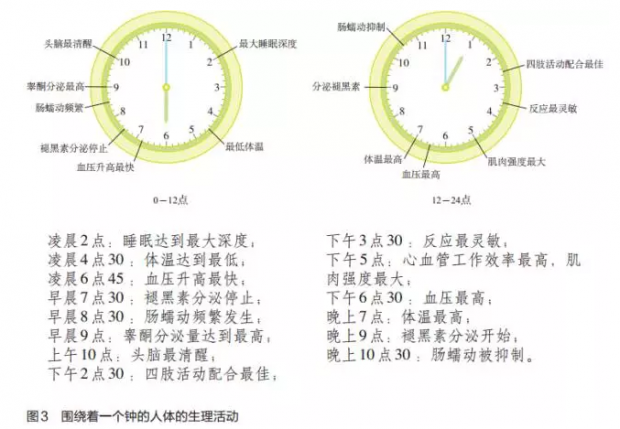
生物钟的正常工作对人的健康起着重要作用。生物钟失调会导致失眠、体乏、抑郁、免疫功能低下甚至产生包括肿瘤在内的各种疾病。根据人的生理生化活动的周期性变化,人可以合理安排一天的活动,从而使工作和休息效率达到最高,也使得人的身心健康状态达到最佳。
因此,钟是一个计时器和报时器,它记录和告知我们事件在时间上的规律性。
钟是一种力量,它指挥着世界万物按不同的周期,有序地做节奏性运动。生物钟是大自然中各种自然钟的一种,是生命物质适应物质世界基本运动规律的一种生命运动形式,是大自然对生物演化的选择,它赋予了生命以预见和应对自然环境变化的能力。了解和顺应大自然赋予我们的生物钟,会使我们的生活更加健康、和谐和愉悦。
作者简介

注:本文节选自《饶有兴趣的大脑》一书。
附作者参与这项工作的主要文献
1. Q. Yu, A. G. Jacquier, Y. Citri, M. Hamblen, J. C. Hall and M. Rosbash. (1987). Molecular mapping of point mutations in the period gene that stop or speed up biological clocks in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 84: 784-788.
2. Q. Yu, H. V. Colot, C. P. Kyriacou, J. C. Hall and M. Rosbash. (1987). Behavior modification by in vitro mutagenesis of a variable region within the period gene of Drosophila. Nature 326: 765-769.
3. Y. Citri, H. V. Colot, A. C. Jacquier, Q. Yu, J. C. Hall, D. Baltimore and M. Rosbash. (1987). A family of unusually spliced and biologically active transcripts is encoded by a Drosophila clock gene. Nature 326: 42-47.
4. Liu, X., L. Lorenz, Q. Yu, J. C. Hall, and M. Rosbash. (1988). Spatial and temporal expression of the period gene in Drosophila melanogaster. Genes & Dev. 2: 228-238.
5. J. C. Hall, S. J. Kulkarni, P. Kyriacou, Q. Yu and M. Rosbash. (1990). Genetic and molecular analysis of neural development and behavior in Drosophila. In: Developmental Behavior Genetics (ed. M. E. Hann), pp100-112, Oxford Univ. Press, New York.
6. Liu, X., Q. Yu, Z., Huang, L. J. Zweibel, J. C. Hall, and M. Rosbash. (1991). The influence of altered spatial pattern of period gene expression in the strength and period of circadian clock in Drosophila melanogaster. Neuron 6: 753-766.
7. Wheeler, D. A., C. P. Kyriacou, M. L. Greenacre, Q. Yu, J. E. Rutila, M. Rosbash, and J. C. Hall. (1991). Molecular transfer of a species-specific courtship behavior from Drosophila simulans to D. melanogaster. Science 251: 1082-1085.

话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}