阅读:0
听报道
撰文|徐小冬(河北师范大学生命科学学院、中国细胞生物学学会生物节律分会副会长)
责编|叶水送
● ● ●
2017年10月2日,诺贝尔生理或医学奖授予发现调控果蝇昼夜节律分子机制的三位美国科学家。当晚,笔者的同行们在微信上欢欣鼓舞时,自家阳台上那颗仙人球正处于每年如约而至的花期,并且在晚上10点左右准时怒放,优雅如凌波仙子。此刻在遥远的北美大陆,成千上万的橙色斑纹与黑色翅脉相间的帝王蝶,正在从北美洲长途迁徙到温暖的墨西哥冷杉林中过冬的路上。

“天地革而四时成”——季节节律
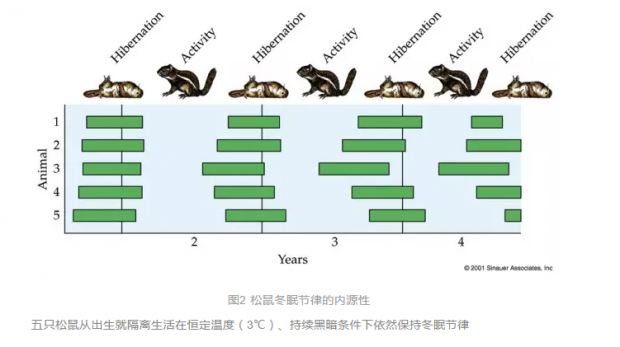
上述提到的植物季节性开花和帝王蝶迁徙都属于生物对季节变化的响应,自然界中季节节律的例子比比皆是,如植物季节性开花结实、动物季节性休眠、迁徙、繁殖,人类季节性情感失调等。内源性是昼夜节律的特点之一,那么季节节律或者说近年节律又如何呢?一个非常有趣的实验表明,把刚出生的松鼠,隔离培养在黑暗、3℃恒温条件下,松鼠的冬眠依然表现出近年节律,这似乎说明松鼠冬眠的季节节律是内源性的,以及可被遗传(图2)。
上面的例子容易使人想到一个问题:与生活在四季分明的高纬度地区的生物相比,赤道附近的热带地区的动植物也具有年节律性吗?
另一个经典的季节节律的研究是以热带的野鹟为研究对象,野鹟生活在光温周期相对恒定的肯尼亚地区,其繁殖和换羽时间也表现为年节律;将雏鸟从肯尼亚转移到德国,在恒定的温度和光照条件下抚养,它们的睾丸宽度(季节性繁殖指标)和换羽时间依然表现出很强的年节律性。结合季节节律的相关研究,时间生物学领域的奠基人之一的尤金·阿绍夫提出,外界信号是作为与环境同步化的“授时因子”,通过影响机体内源的生物钟起作用,而不是通过直接影响(诱导或抑制)机体内部的生理活动。

自带有生物钟校准功能的GPS——帝王蝶的漫漫迁徙路
帝王蝶是目前认为研究较为清楚的进行季节性长途迁徙的蝴蝶。这种具有传奇色彩的蝴蝶的迁徙和鸟类迁徙有所不同,一般来说北美落基山脉以东的帝王蝶完成一次迁徙至少需要三代,新生蝴蝶在没有亲属引领下,前仆后继才能完成。
史蒂芬·瑞普尔特就是研究这一传奇现象的科学家之一。他给人印象最深的就是一头爱因斯坦式蓬松的白发,和他交流收获的不仅是科学上启迪,也能深深感到他对科研发自内心的热爱,一个科学家觉得科研“很好玩”并作为终生职业,这应该是科学研究的最高境界吧。
瑞普尔特教授从小就对鳞翅目昆虫——惜古比天蚕蛾着迷,在他的职业生涯中还发表过非常出色的关于这种蛾子的生物钟相关研究论文。

瑞普尔特博士和合作者长期以来从生理生化水平到分子水平,通过巧妙的实验设计和艰辛的工作,逐步揭示了和帝王蝶迁徙相关的复杂调控机制,特别是近些年完成了帝王蝶基因组测序工作,并将基因编辑技术应用于相关研究,从系统和进化角度进一步开展了广泛和深入的研究(图4)。
和我们自驾游需要GPS一样,帝王蝶长途迁徙也需要一套GPS系统,它的GPS系统非常高端,是时间补偿性的太阳罗盘,通过眼部视觉系统感受“天空光”的信号(Skylight cues,包括蓝紫色、蓝色和绿色的梯度变化,太阳的位置以及紫外光的偏振模式),因为“天空光的信息是随时间变化的,所以必须同时有一套时间补偿机制共同起作用才能完成较为精准的定位。研究者揭示了帝王蝶的两个触角里面的生物钟可为飞行定位提供时间补偿,最终两类信号在脑部中央复合体和相关神经节进行整合,再把信号传递给运动系统从而完成定位飞行。帝王蝶生物钟的分子机制和果蝇类似,是由PER、TIM、CRY2、CLK、CYC、VRI和PDP1等组分构成的多层次不同水平上的转录/翻译反馈环路和调节环路。
“用一朵花开的时间”迷上生物钟——昼夜与季节节律的完美结合
不论多年生植物还是一年生植物都会响应季节的变化。生物钟输入途径中两个最重要的授时因子是光照和温度(还包括营养等其它因素),而季节的周期性也主要是这两个因素的变化。动植物的季节性生理活动周期一定程度上依赖于日照长短的规律性变化。上世纪20年代,怀特曼·加纳和亨利· 阿拉德提出光周期和光周期现象的概念,并根据植物开花对光周期中日照长度的要求,将植物分为长日照植物(小麦、莴苣等)、短日照植物(大豆、水稻、菊花等)和日中性植物(番茄、月季等);而后续研究中对暗间断处理实验结果表明,植物是通过感知周期中黑暗的时长来对光周期做出响应的。植物对光周期的响应除了开花,还包括块茎的形成、芽的休眠等。
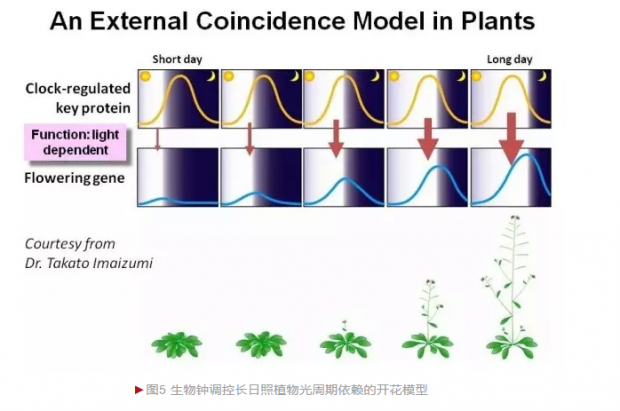
在自然界,随着外部环境信号的季节性轮回,植物的生长发育每天都发生一定的小幅度变化,这种变化每天都在“校准”内源性的生物钟,提高对环境适应的精度。对于长日照或短日照植物的开花来讲,生物钟调控的光周期依赖途径至关重要:通过生物钟输入途径感受外部环境的光周期变化,进而改变生物钟核心组分的表达,并通过一系列信号转导途径影响到FT基因(FT蛋白被认为可能是人们长久以来寻找的“成花素”——Florigen组分)的表达,从而促进或抑制开花。
光周期途径的调控机制是多层次、不同水平上发生的,不仅包括了转录和转录后的mRNA拼接,翻译和翻译后蛋白修饰及降解等途径,还涉及到空间因素(比如FT蛋白从叶片转移到顶端分生组织)和时间因素(比如FKF1和GI在短日照和长日照中的昼夜时间相位是否相同,影响到其蛋白复合体的形成,进而决定了是否能够解除CDF1对CO的抑制,从而影响到CO对FT的激活)。组学分析结果表明几乎所有光周期开花途径的相关基因或蛋白的昼夜节律性表达都处在生物钟调控之下,从时空动态调控上看,生物钟决定的开花相关各组分节律性出现的相位是一个极其重要的因素(图5)。
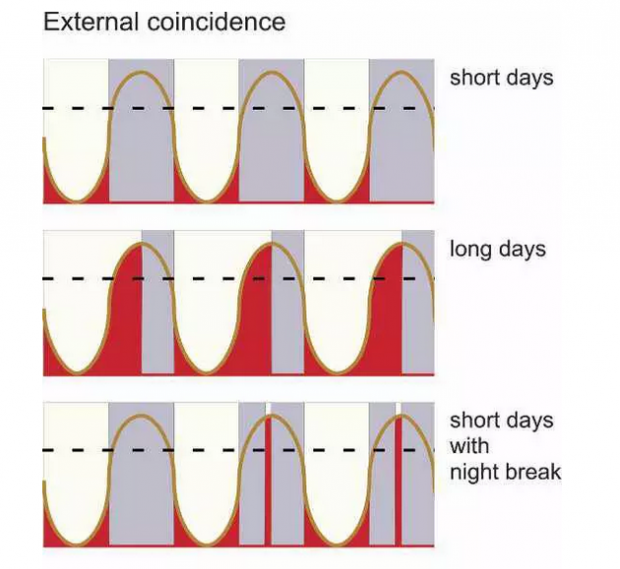
1932年,欧文·邦宁博士用植物的遗传学杂交实验证实了周期节律的可遗传性;1936年,他又根据对生物钟与大豆的光周期响应的研究首次提出了如下假说来解释光周期现象:一天中的24小时可分为喜光区段(对光敏感)和厌光区段(对黑暗敏感),两种区段的节律性位移由内源生物钟调控;外部环境中的光信号出现在上述两个区段与否的信息使得植物感知或判断自身是处于长日照条件下或短日照条件下。
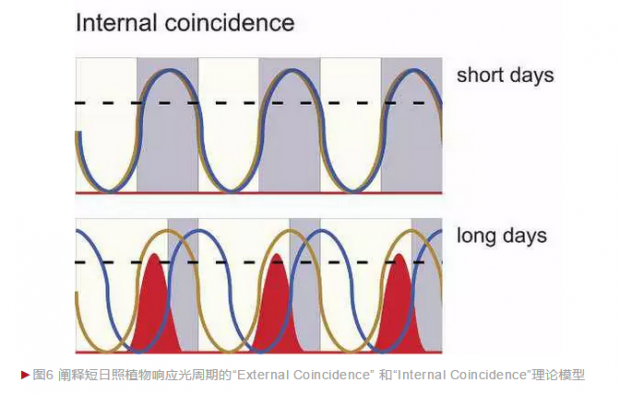
时间生物学领域的奠基人之一科林·布里斯顿进一步发展了 邦宁的假说,确立了“External Coincidence” 、“Internal Coincidence”理论模型:其中光信号起到两个关键的作用,一是设置生物钟使得开花相关因子的节律性表达峰值出现在下午(与长日照条件下光信号出现的时间一致/External Coincidence),二是调节开花关键组分的活性。长日照条件下,光通过生物钟调节的开花关键组分作为正调因子促进植物的开花;而短日照条件下,光调节的该组分作为开花调控的抑制因子起作用(图6)。
介导动植物季节响应的除了光周期之外,温度也是重要的授时因子,例如,春化过程中的低温信号也是调控植物季节性开花的重要因素。生物钟基因也会对低温信号做出响应,例如漫长而寒冷的冬季过后,杨树生物钟基因PttLHY的表达有利于打破侧芽的休眠,使之在早春适时萌发出新的枝条。
从生到死,植物的一生的生长发育都离不开生物钟调控。以模式植物拟南芥为例,一粒种子吸涨后起始萌发,生物钟就开始启动,最有意思的是即使去除外界光照、温度的影响,比如把种子从一开始就在播种在恒温持续黑暗或恒温持续光照的条件下,生物钟基因的转录依然会产生近24小时的振荡模式,具体是什么信号引导了生物钟启动还需进一步研究。研究表明植物叶片衰老时生物钟会变快,但目前生物钟变化是决定衰老的原因还是结果尚有争论。
亚日节律(一般指周期大于28小时的节律)除了季节节律,还包括近周节律、近月节律、近年节律和近十年节律等;与之对应的还有日内节律/超日节律(一般指周期小于20小时的节律常见的节律现象)。
近潮汐节律也是一类比较有趣的节律现象,一些海岸线的生物,例如水生的蟹类动物有这种节律,近潮汐节律的调控也是内源的,不依赖与潮汐等外界环境。
总之,生物钟节律与其它生物节律的关系依然有待于进一步研究,从进化上讲,内源节律的精确计时,使得机体提前“预知”环境的周期性变化并与之同步,增强生物对环境的适应性。
译名对照表:
帝王蝶 Danaus plexippus
季节节律 Seasonal rhythm 或 Circaseasonal
季节性情感失调 SAD, Seasonal Affective Disorder
近年节律 Circannual
松鼠 Golden-mantled ground squirrels
野鹟 African stonechat
尤金·阿绍夫 Jürgen Aschoff
“授时因子” Zeitgeber,德语,Time giver
史蒂芬·瑞普尔特 Steven M. Reppert
惜古比天蚕蛾 Hyalophora cecropia,北美洲最大的蛾子之一
转录/翻译反馈环路 TTFL, transcriptional/translational feedback loop
怀特曼·加纳 Wightman Garner
亨利· 阿拉德 Henry Allard
光周期 Photoperiod
光周期现象 Photoperiodism
暗间断处理 Night break
黑暗的时长 Critical Night Length
欧文·邦宁 Erwin Bünning
喜光区段 photophile phase
厌光区段 scotophile phase
科林·布里斯顿 Colin Pittendrigh
春化 Vernalization
亚日节律 Infradian rhythm
日内节律/超日节律 Ultradian
近潮汐节律 Circatidal
参考文献
[1] IMAIZUMI T, SCHULTZ T F, HARMON F G, et al. FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis [J]. Science, 2005, 309(5732): 293-7.
[2] REPPERT S M. A colorful model of the circadian clock [J]. Cell, 2006, 124(2): 233-6.
[3] ZHAN S, MERLIN C, BOORE J L, et al. The monarch butterfly genome yields insights into long-distance migration [J]. Cell, 2011, 147(5): 1171-85.
[4] SONG Y H, SHIM J S, KINMONTH-SCHULTZ H A, et al. Photoperiodic flowering: time measurement mechanisms in leaves [J]. Annu Rev Plant Biol, 2015, 66: 441-64.
[5] GWINNER E. Circannual rhythms in birds [J]. Curr Opin Neurobiol, 2003, 13(6): 770-8.
[6] KOBAYASHI Y, WEIGEL D. Move on up, it's time for change--mobile signals controlling photoperiod-dependent flowering [J]. Genes Dev, 2007, 21(19): 2371-84.
[7] REPPERT S M, GUERRA P A, MERLIN C. Neurobiology of Monarch Butterfly Migration [J]. Annu Rev Entomol, 2016, 61: 25-42.

话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}