阅读:0
听报道
撰文 | 王承志
责编 | 陈晓雪
细胞对于外界信号的感知能力是基本的生物学需求,其中很重要的一种是对机械力的感知。比如听觉和触觉,肺对扩张的感知,血管壁对血流的感知,许多生理过程都与机械力的感知有关。一些特化的细胞进化成为专门感知机械力的“终端”,比如内耳毛细胞、本体感觉神经元等等,它们通常对机械力非常敏感。不仅如此,大部分细胞都含有感知机械力的结构,这对于细胞迁移、扩增以及分化等功能都很重要。
细胞对机械力的“感知”需要通过把机械力转化为细胞能够识别的生物信号,这通常是由离子通道和细胞膜受体以及细胞内信号传导通路共同实现。早在1979年,加州理工学院的Hudspeth(现为洛克菲勒大学教授)和Corey(后在哈佛大学任职),就通过在牛蛙听觉上皮细胞中的实验,推测机械力刺激可能通过离子产生的电流传导,因为亚毫秒级别的信号传导不可能通过传统的第二信使的方式传递[1]。2010年,美国斯克里普斯研究所Ardem Patapoutian实验室发现了高等动物中,Piezo1和Piezo2 两个基因是编码哺乳动物机械门控阳离子通道的必要组成成分[2]。

后来的研究发现,Piezo1和Piezo2属于一类很保守的机械门控阳离子通道,在包括触觉、痛觉、呼吸、本体感受和血管发育等多种生理过程中起作用(图1)。人体中这些基因突变会导致红细胞干瘪综合症、远端关节扭曲症、先天性淋巴管发育不良和触觉缺失等疾病,显示出它们的重要性[3]。
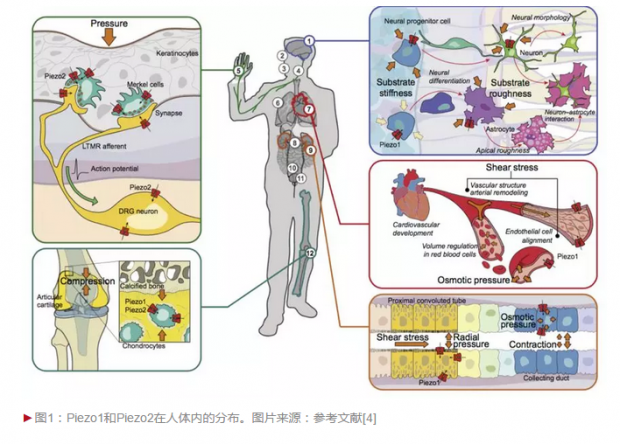
从预测的结构看,这类蛋白与其它所有已知蛋白都没有多少相似性,而且相比其它离子通道,这类蛋白显得非常巨大(超过2000个氨基酸)。清华大学药学院研究员肖百龙博士在Ardem Patapoutian教授实验室从事博士后期间与其同事首次证明Piezo蛋白形成机械门控阳离子通道,这在哺乳动物细胞中是首次被鉴定发现的,但其具体的作用机制并不清楚[6]。
蛋白质的三维结构通常能够帮助科学家解释其功能。2015年,清华大学高宁课题组、肖百龙课题组与杨茂君课题组合作在《自然》杂志报道了Piezo1通道中等分辨率的冷冻电镜三维结构,揭示了其三叶螺旋桨状的三维构造特征,为解析其分子作用机制提供了重要线索[5]。
2016年,肖百龙课题组在《神经元》杂志发表文章,报道了Piezo1通道负责离子通透与选择性的孔道区模块以及负责机械力感受与传导的机械传感模块的氨基酸构成,进而首次提出Piezo1通道以功能区模块化的方式来行使其机械门控离子通道功能的假说[7]。
2017年,肖百龙课题组在《自然-通讯》杂志进一步报道了Piezo通道的新型调控蛋白SERCA2,并系统阐明了其对Piezo蛋白通道活性调控的作用机制,为基于这一蛋白互做调控模式的潜在药物开发提供了重要线索[8]。
本周二,《自然》在线发表了清华大学肖百龙与李雪明课题组题为“Piezo1离子通道的结构与机械门控机制”(Structure and Mechanogating Mechanism of the Piezo1 Channel)的研究长文[9]。该研究解析了哺乳动物机械门控Piezo1离子通道的高分辨率三维结构,揭示了其参与机械力感受与传递的关键功能位点,进而首次提出了Piezo通道以类似杠杆原理进行机械门控的精巧工作机制。值得一提的是,最初发现Piezo1和Piezo2基因的Ardem Patapoutian实验室也和同在斯克里普斯研究所的Andrew Ward实验室合作,几乎同时在《自然》杂志上发表了Piezo1的高分辨率结构[10]。洛克菲勒大学的Roderick MacKinnon实验室也同时在eLife期刊发表了Piezo1的高分辨率结构[11]。
Piezo1通道整体呈现三聚体三叶螺旋桨状结构(图2a),其羧基端约350个氨基酸组成中心的控制离子通透的中心孔道部分,其氨基端的2200个氨基酸组成外周的特征性结构域,包括“桨叶”(blade)、“长杆”(beam)以及“锚定区”(anchor)(图2a-c)。
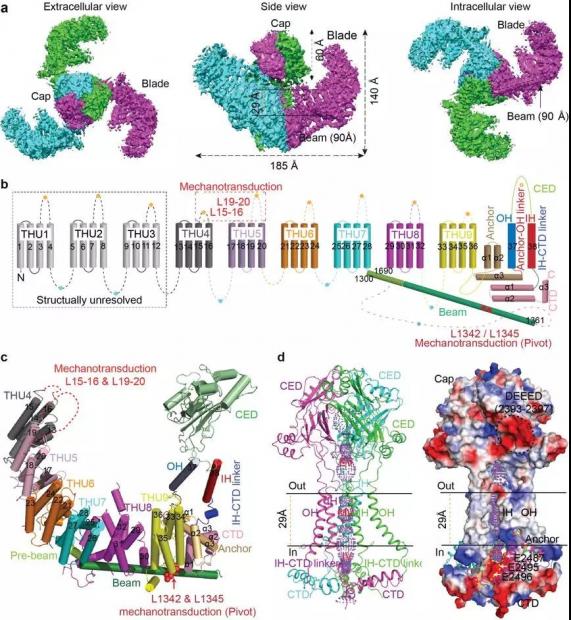
►图2:Piezo1三维结构。图片来源:参考文献[9]
虽然Piezo1蛋白并不包含重复性序列,肖百龙等学者却惊奇地发现其外周桨叶部分由共9个重复性的、以4次跨膜区为基础的结构单元串联而成。他们把这一特征性的结构域命名为Transmembrane Helical Unit (THU),这也恰好是清华大学的英文缩写。而在Ardem Patapoutian和Andrew Ward的论文中,这种结构被命名为Piezo repeats。在拓扑结构上,Piezo1蛋白以每个亚基包含38次跨膜区、总计114次跨膜区的形式,组装成目前已知的跨膜次数最多的一类膜蛋白复合物(图2b-c)。
在三维结构的胞内部分,存在着长约90 Å的特征性beam结构,其将远端桨叶区连接到中心孔道区部分的CTD以及anchor结构域(图2b-c)。肖百龙认为这是Piezo通道将外周桨叶部分所感知的机械力变化有效传递到中心孔道部分的重要结构基础。结构显示beam区存在着不均一的相对位移,即远端的运动位移更大,而靠近中央成孔区只显示轻微的位移变化。因此,Piezo1结构的构象变化契合了杠杆作用原理。他们的电生理功能研究数据进一步支持了其提出的Piezo1通道以杠杆原理进行机械力感知的机制。
肖百龙博士告诉《知识分子》,以上研究成果有力地推动了对Piezo通道结构特征与分子机制的理解。他们还发现Piezo诸多与人类遗传疾病(例如Piezo1、Piezo2基因突变分别导致红细胞干瘪综合症以及远端关节扭曲症)相关的氨基酸突变,集中在beam-CTD-Anchor的相互作用界面,为下一步深入理解这些遗传疾病突变如何导致Piezo通道功能失常提供了重要线索。
参考文献:
1. Corey DP and Hudspeth AJ: Response latency of vertebrate hair cells. Biophys J 26:499-506, 1979.
2. Coste B, Mathur J, Schmidt M, et al.: Piezo1and Piezo2 are essential components of distinct mechanically activated cationchannels. Science 330: 55-60, 2010.
3. Murthy SE, Dubin AE and Patapoutian A: Piezosthrive under pressure: mechanically activated ion channels in health anddisease. Nat Rev Mol Cell Biol 18: 771-783, 2017.
4. Wu J, Lewis AH and Grandl J: Touch, Tension,and Transduction - The Function and Regulation of Piezo Ion Channels. Trends BiochemSci 42: 57-71, 2017.
5. Ge J, Li W, Zhao Q, et al.: Architecture ofthe mammalian mechanosensitive Piezo1 channel. Nature 527: 64-69, 2015.
6. Coste B, Xiao B, et al.: Piezo Proteins Are Pore-forming Subunits of Mechanically Activated Channels. Nature. 2012.
7. Zhao Q, Wu K, Geng J, et al.: Ion Permeationand Mechanotransduction Mechanisms of Mechanosensitive Piezo Channels. Neuron 89: 1248-1263, 2016.
8. Zhang T, Chi S, Jiang F, Zhao Q and Xiao B:A protein interaction mechanism for suppressing the mechanosensitive Piezochannels. Nat Commun 8: 1797, 2017.
9. Structure and Mechanogating Mechanism of the Piezo1 Channel,
DOI: 10.1038/nature25743
10. Saotome K, Murthy SE, Kefauver JM, WhitwamT, Patapoutian A and Ward AB: Structure of the mechanically activated ionchannel Piezo1. Nature 2017.
11. Guo YR and MacKinnon R: Structure-based membrane dome mechanism for Piezo mechanosensitivity. Elife 2017.

话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}