阅读:0
听报道

美国迈克尔·希茨,现为德克萨斯大学加尔维斯顿医学分部终身教授,此前为新加坡国立大学生物力学研究所所长。图源:
前 言
如果把一个神经元细胞想成中国地图的话,那么将物品从广州运到内蒙古的 “快递小哥” 是谁呢?迈克尔·希茨和罗纳德·韦尔试图在乌贼身上寻找答案,然而那一年,由于全球经历了严重的厄尔尼诺现象,美国西海岸的乌贼全溜了……
“直到今天,不断地迎接意料之外的实验结果的挑战依然令我感到兴奋,并且是推动我在科学事业上不断进取的最大动力。”
—— 迈克尔· 希茨
撰文 | 黄宇翔
责编 | 万 朵
结缘生物化学研究(1946-1972)
1946年12月,刚刚从美国海军退役的大卫· 希茨(David Sheetz)迎来了自己第一个孩子的降生,为他取名为迈克尔· 希茨(Michael Sheetz)。退役后的大卫在内布拉斯加大学取得化学博士学位,并在大名鼎鼎的陶氏化学公司的研发部门找到了一份工作,随后举家搬到陶氏公司的总部——位于美国密歇根州的米德兰(Midland)。[2]
迈克尔在米德兰度过了他快乐的童年时光。这座只有四万人口的小城,有着浓郁的化学氛围——化学家赫伯特· 陶(Herbert Dow)1897年在这座小城创立了陶氏公司。这家最初以经营漂白剂和溴化钾为主要业务的公司在二十世纪上半叶蓬勃发展,到了五十年代已经是一家年销售额达10亿美元的化工巨头。由于大量米德兰的居民都是陶氏公司的雇员,因此这座小城也被称作 “陶氏化工之城”(A Dow Chemical Town)。
“大家都期望陶氏公司化学家们的孩子在未来都能成长为比他们父辈更为优秀的化学家,我当时深受这种思想的影响。” 迈克尔回忆自己的童年时说。[3]
1964年,高中毕业的迈克尔选择到离家140英里的阿尔比恩文理学院(Albion College)就读。这家文理学院的化学系不错,而且在迈克尔看来更重要的是 “在这里我能获得更多老师一对一的指导,能有机会做自己独立的项目”。[3]
在阿尔比恩文理学院,迈克尔打下了扎实的数理基础,并且对基础物理学和化学研究产生了浓厚的兴趣,但身为化学家的父亲大卫希望大儿子能打开视野。
“老爸尝试说服我能治病救人的研究比物理或者化学的基础理论更有价值,” 迈克尔在2012年拉斯克奖颁奖典礼的致辞中回忆道,“为了验证他的观点,我去医院做了几个月的夜间护工,很快就意识到在当时许多治疗方法都遵循着 ‘不造成伤害’(do no harm)的原则,根源是我们那时对人体的生物化学机制了解还非常匮乏。”
1967年,在参加了美国阿贡国家实验室(Argonne National Laboratory)暑期科研项目后,迈克尔正式将自己的志向定为成为一名生物化学家,并于1968年被加州理工学院研究生项目录取。
在加州理工学院,迈克尔在生物物理化学家 Sunney Chan(陈长谦)的指导下用核磁共振影像方法对细胞膜的性质进行研究。然而,受于技术手段的局限,迈克尔在研究生期间仅仅能在相对剧烈的条件下(如高温变性)检测到细胞膜的核磁共振信号,而无法对细胞膜更细微的结构进行观测。[4-6]
1972年,迈克尔获得博士学位,前往加州大学圣迭戈分校(UCSD)的 Jon Singer 实验室做博后研究。
(注:父亲大卫的引导不仅影响大儿子迈克尔走向生物化学研究的道路。迈克尔的弟弟马修· 希茨(Matthew Sheetz)在化学家父亲的影响下进入生物医学领域,并在礼来公司作为临床研究医师领导包括针对糖尿病在内的超过20项临床试验。)
初出茅庐(1972-1982)
在迈克尔来到 Jon Singer 实验室的70年代,人们关于细胞膜生物化学的知识还非常缺乏。迈克尔和导师 Jon Singer 针对 “细胞膜双分子结构中内膜富含阴离子脂质” 的这一事实提出了一个他们称之为 “双层偶联”(bilayer couple)的假设:阴离子内膜会吸引一些阳离子药物,进而通过影响膜结构改变细胞的形状。通过简洁的功能学和电镜实验,迈克尔作出了自己在学术界第一个重要的贡献,推动了 “双层偶联” 模型的发展。[7]
1974年,迈克尔在康涅狄格大学建立了自己的独立实验室,继续自己博后期间在红细胞细胞膜方面的工作,试图探究细胞感受 “双层偶联” 的生物化学机制。红细胞细胞膜的膜蛋白扩散的速度比其他细胞类型细胞膜膜蛋白扩散速度要慢约100倍,而迈克尔和他在康涅狄格大学的合作者 Dennis Koppel 和 Mel Schindler 发现,珠蛋白(一种膜骨架蛋白)突变的小鼠的红细胞膜蛋白扩散速度与其他细胞的膜蛋白一样快。基于此,他们提出珠蛋白在细胞膜表面形成局部的 “珊瑚样结构”,能允许膜蛋白局部的扩散,但却阻止其长距离扩散。这一发现为迈克尔带来了他在《自然》杂志发表的第一篇论文,也让他在学术界开始小有名气。[8]
这个时候,迈克尔选择在斯坦福大学的詹姆斯·斯普迪赫(James Spudich)实验室开启一段改变了他职业生涯的学术休假。
黏菌,丽藻,鱿鱼,厄尔尼诺以及驱动蛋白的发现(1982-1985)
当时詹姆斯·斯普迪赫希望能通过体外重组得到肌动蛋白运动的过程。
詹姆斯仅仅比迈克尔年长四岁,在一个克罗地亚移民家庭长大,研究生期间在斯坦福大学 Arthur Kornberg(1959年因发现DNA聚合酶获诺贝尔生理医学奖)实验室接受了严格的生物化学训练。博士毕业后,他留在斯坦福大学 Charley Yanofsky 实验室做了一段短期博后,运用遗传学手段研究大肠杆菌色氨酸操作子,随后来到英国剑桥大学的 Hugh Huxley 实验室研究肌动蛋白运动的机制。Hugh Huxley 基于X射线衍射与电镜实验结果为提出解释肌肉如何收缩舒张的 “肌丝滑动” 模型作出了重要贡献。詹姆斯在 Hugh Huxley 实验室的博后工作提出原肌球蛋白(tropomyosin)和肌钙蛋白(Troponin)通过空间位阻抑制肌丝蛋白(actin)与肌动蛋白(myosin)的相互结合。[9-12]
1984年,James Spudich在显微镜前
在1970年代,对于肌丝滑动模型最大的挑战是由于细胞内存在各种可能与肌丝蛋白结合的蛋白,研究者们无法清晰地阐明 ATP 水解引发机械运动的分子机制。因此,在加州大学旧金山分校(UCSF)建立独立实验室的詹姆斯希望能在体外重现肌丝蛋白与肌动蛋白的相互作用的生理过程(詹姆斯后于1977年搬到了斯坦福大学结构生物学系)。
为了实现这个目标,詹姆斯率领团队在十年的时间里尝试了包含粗糙脉孢菌(Neurospora crassa)、酿酒酵母(Saccharomyces cerevisiae)、黏菌(Physarum polycephalum)、盘基网柄菌(Dictyostelium discoideum)等在内的许多体系。在最初的尝试中,黏菌是最被詹姆斯寄予厚望的实验体系,詹姆斯的博后 Margaret Clarke 成功从黏菌中纯化出了肌动蛋白,并且黏菌的肌丝蛋白与细胞膜紧密相连,使得研究者可以相对容易地分离出结合聚苯乙烯珠子的肌丝蛋白。詹姆斯团队试图通过珠子将肌丝蛋白固定在玻璃上,然后向其中加入结合了肌动蛋白的珠子和提供能量的 ATP,希望能观测到肌动蛋白-珠子在肌丝蛋白上的定向移动。[13-15]
遗憾的是,直到1982年学术休假的迈克尔加入其团队时,詹姆斯依然没有成功建立起这一体外重组体系。
人们事后诸葛亮分析,詹姆斯团队前期失败的原因很可能是固定在玻璃板上的肌丝蛋白朝向各异,导致只能观察到肌动蛋白-珠子的一些无规则的运动。
迈克尔尝试利用丽藻(Nitella axillaris)建立詹姆斯团队长期希望实现的体外重组体系。
结构生物学家施一公在给清华生科院的本科生上课时曾经说过,实验所选用的体系之于生物学研究,正如引擎之于汽车,体系如果用对了,往往能达到事半功倍的效果。
对于建立体外肌丝滑动模型这一特定的生物学问题,迈克尔幸运地选择了合适的实验体系:丽藻的大节间细胞(giant internodal cell),叶绿体紧紧贴在细胞壁上,成排的肌丝蛋白整齐地依附在叶绿体上(每排叶绿体平均含有5排肌丝蛋白),而每排光学显微镜下看到的肌丝蛋白丝含有上百个肌丝蛋白分子,排列得宛如阅兵方队一般整齐,仿佛只等待着研究者去检阅。
通过显微镜,迈克尔见证了这一细胞内的生命奇观,并将其应用于回答自己的科学问题上。神奇至极,这一次,詹姆斯团队第一次在体外体系看到了肌动蛋白-珠子在肌丝蛋白的定向移动![16]
1983年5月5日,迈克尔和詹姆斯在《自然》杂志与全世界分享他们成功建立出重组肌丝滑动体系的喜悦。
这场科学的胜利,丽藻功不可没。
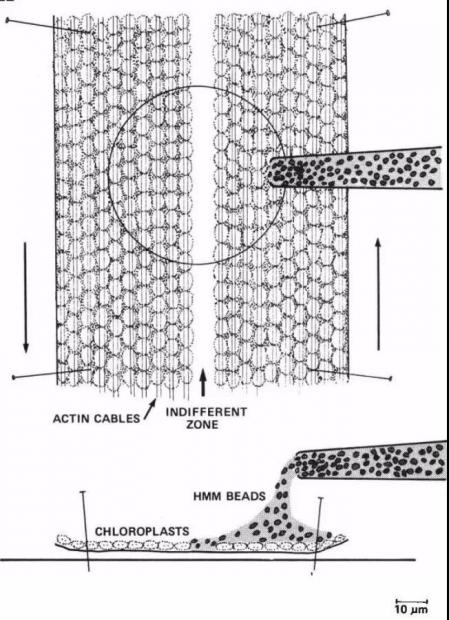
丽藻节间细胞:完美的体系!来源:Ref.16
1982年迈克尔利用丽藻成功建立体外重组体系的消息,震动了他们实验室楼下的一位23岁的研究生,罗纳德·韦尔(Ronald Vale)。
罗纳德此时在 Eric Shooter 教授的指导下研究神经生长因子如何引起神经生长因子受体激活的生物化学机制。他在研究中不禁思考:对于神经元而言,在树突附近的信号激活是如何将特定的物质从胞体(神经元的细胞核) “远行千里” (对于细胞而言的确如此!)运输到轴突的末端呢?如果把一个神经元细胞想成中国地图的话,那么将物品从内蒙古运到广州的“快递员”是谁呢? [17,18]
罗纳德立刻想到,迈克尔利用丽藻细胞中揭示的机制在神经元中同样适用——即神经元中快递运输的 “铁路” 就是肌丝蛋白,肌动蛋白就是细胞中勤勉的 “快递小哥”!
激动的罗纳德立刻找到了楼上的迈克尔,想同他一起揭晓神经元中 “快递传输” 的分子机制。
这一次,他们所选用的研究体系是为现代生理学立下过汗马功劳的乌贼巨大轴突。
这一年罗纳德23岁,迈克尔36岁。下面就以小罗和小麦称呼他俩吧。
天公作美,就在同一年,伍兹霍尔海洋研究所(Woods Hole Marine Biological Laboratory)的 Robert Allen 和 Shinya Inoue 独立地开发出能将显微镜图像呈现在电子屏幕的技术,大大解放了研究者观察记录的负担。并且 Robert Allen 团队用此技术成功地观察到乌贼巨大轴突中显著的轴浆运输过程。[19-21]
经过讨论,小罗和小麦设计了如下实验:他们计划将丽藻模型依葫芦画瓢,将鱿鱼巨大轴突的肌丝蛋白分离出来,加入肌动蛋白-珠子,期望能观察到珠子的定向移动。
怀着激动的心情,小罗向隶属于斯坦福大学海洋实验室的霍普金斯海洋监测站(Hopkins Marine Station)打电话请求他们从海里打捞一些鱿鱼作为实验材料,对方爽快地答应了。
经过亿万年演化形成的胞内物质运输的分子机制,即将在凡人面前被揭晓!
天将降大任于斯人之前,总是要先制造一些波澜。
小罗和小麦就算是天选之子,也不能够例外。
距小罗给霍普金斯海洋监测站打电话几个月时间过去了,他们始终没有收到对方寄来的鱿鱼,也没有收到任何电话通知。
苦苦等米下锅的小罗终于忍不住了,于是在1983年的6月拨通了海洋监测站的电话。
“我几个月前曾经请求过请你们帮忙捕捞一些鱿鱼,请问你们不会是忘记了吧?”
“没有呀!可是我们今年确确实实捕捉不到鱿鱼。”
这到底是怎么回事呢?鱿鱼怎么会突然抓不到了呢?
关注全球气象变化的读者朋友们可能知道,1983年全球经历了严重的厄尔尼诺现象,其中后果之一就是加州西海岸的海洋温度上升,导致鱿鱼离开海岸到温度更低的海域,该年度鱿鱼捕捞量呈现断崖式下跌![22]
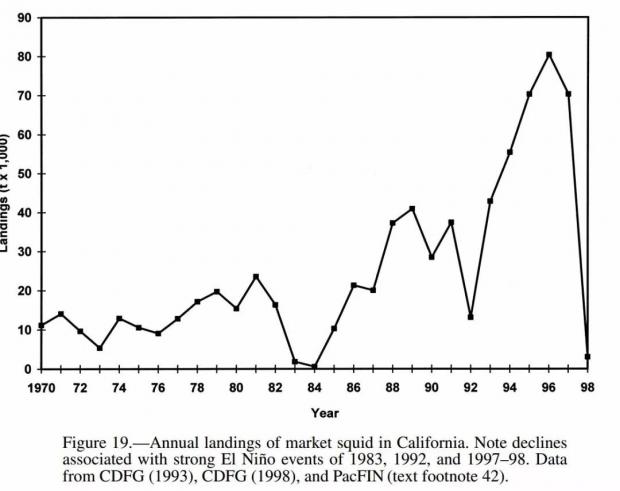
加州鱿鱼捕捞量在1983年由于厄尔尼诺现象呈断崖式下跌。 来源:Ref. 22
巧妇难为无米之炊,没有实验材料,这实验还怎么做呀?
小罗和小麦执着地希望能用实验检测自己的想法。既然加州的西海岸今年捕不到鱿鱼,那就到有鱿鱼的地方去做实验。
于是在1983年的夏天,他俩来到了位于东海岸的伍兹霍尔海洋研究所。
刚刚抵达伍兹霍尔海洋研究所,翻开最新一期《自然》杂志的两人都感到仿佛有一块冰滑入了自己的胃里:英国 MRC 实验室的研究者,报道了几乎与他们计划做的完全相同的实验,只不过所用的实验材料不是乌贼,而是螃蟹。[23]

还没来得及撸起袖子搬砖就被人在《自然》上抢发了?来源:Ref. 23
刚刚千里迢迢从西海岸来到东海岸,还没开始动手就发现自己设计的实验已经被其他人先一步在《自然》杂志抢发了。
小罗和小麦的心情在这一天跌落到了谷底。难道就此收拾行李打道回府吗?
仔细阅读了英国团队的这项研究后,他们发现此研究所使用的珠子与他们实验设计的不同,是不结合肌动蛋白的塑料珠,并且也没有对所观察到的不含有肌动蛋白结合的珠子运动从机制上给出明确的解释。也就是说,现在开始在鱿鱼的体系下用肌动蛋白-珠子做实验开始追赶,依然很有价值。
经过了一番折腾,小罗和小麦终于做了他们在几个月前计划的实验。但实验的结果却令他们大跌眼界:被寄予厚望的肌动蛋白-珠子实验组没有表现出定向的移动,而没有结合肌动蛋白在此处被用作阴性对照的珠子却表现出定向的移动!

令人费解的结果 来源:Ref. 24
面对如此费解的结果,聪明如小罗和小麦也感到一筹莫展。
他们重新回顾了他们的假设模型,仔细思索自己的实验设计哪里可能存在问题。
聪明的读者朋友们也许已经发现了,他们假设自己从从乌贼巨大轴突分离出的纤维束的主要成分是肌丝蛋白,因此预测肌动蛋白-珠子能与之结合进而在纤维束上作定向运动。但假若这些纤维束的成分不是肌丝蛋白呢?
伍兹霍尔海洋研究所的 Bruce Schnapp 和 Thomas Reese 给予了小罗和小麦很大的帮助。为了鉴定小罗和小麦分离出的纤维束是否为肌丝蛋白,他们进行了电镜成像实验。实验结果清晰地显示——这些纤维束根本不是肌丝蛋白的结构,预期相反,其成分很可能是微管蛋白![25]
这一结果令小罗和小麦激动异常:纤维束是微管蛋白!这是否意味着在鱿鱼轴浆运输中存在一套全新的分子机制?
他们在逐步走近自然的真相。
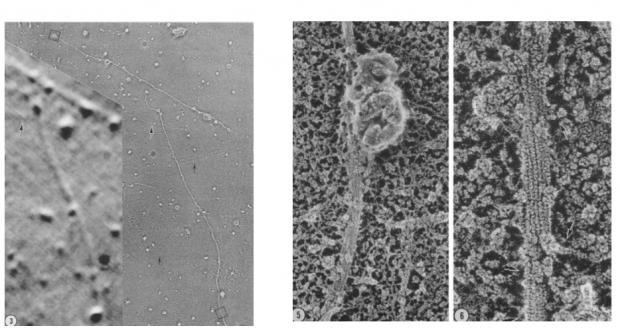
小罗和小麦从乌贼巨大轴突分离出的“肌丝蛋白”未必是肌丝蛋白,而实际是微管蛋白。来源:Ref. 25

小麦,Thomas Reese,Bruce Schnapp和长发飘飘的小罗。来源:Ref. 26
1983年的8月,小罗和小麦试图在体外重组出微管蛋白介导的轴浆运输过程。
他们将纯化出的乌贼的微管蛋白、细胞器组分、ATP 加在一起,没有观察到轴浆运输;
他们将纯化出的乌贼的微管蛋白、细胞器组分、ATP、细胞质基质中的水溶性组分加在一起——奇迹发生了,细胞器囊泡在玻璃片上有序地移动起来![27]
这意味着,细胞质基质水溶性组分中包含一种与肌动蛋白功能类似但是全新的动力蛋白。
接下来的工作重点,就是希望用生化的手段从细胞质基质水溶性组分中包含的上百个蛋白分子中找出那一个全新的动力蛋白。
伍兹霍尔海洋研究所的夏天熙熙攘攘,十分热闹。到了冬天,这里就变得如同修道院一般肃穆寂寥。
1983-1984年的冬天,就在安静的伍兹霍尔海洋研究所,乌贼巨大轴突的细胞质基质水溶性组分通过了一个个柱子被逐步分离成单一组分。终于,在经过一个羟基磷灰石层析柱的洗脱之后,能引发细胞器在微管蛋白上运动的纯净组分终于现出了庐山真面目:聚丙烯酰胺电泳图上清晰的两个条带:大的在110kDa附近,小的在65kDa附近。[28]
千呼万唤始出来,驱动蛋白(Kinesin)就此从人类知识盲区的迷雾中露出了它的神秘面庞。

2012年,迈克尔· 希茨、詹姆斯·普迪赫和罗纳德·韦尔凭借他们对于分子动力蛋白的工作被授予拉斯克基础医学奖。
12年以后,已经是 UCSF 药理系主任的罗纳德解析了驱动蛋白的晶体结构:包含两条重链和两条轻链。重链对应的就是他当初在电泳胶图上看到的100kDa大小的条带;轻链对应的则是那条65kDa左右的条带。[29]
驱动蛋白后续被研究者发现是一类非常大的蛋白超家族,在众多细胞生物学事件和疾病过程中发挥关键作用。[30]
2006年,驱动蛋白这位细胞中的 “快递小哥” 拖动细胞中的 “快递物件” 的视频被哈佛大学的研究者做成了生动的动画,成为了现代细胞生物学的标志。
笔者在高一时第一次看到这个视频,立刻被细胞内部的美所震惊,对生物学研究兴趣大增,如今在接受细胞生物学博士研究生的科研训练。
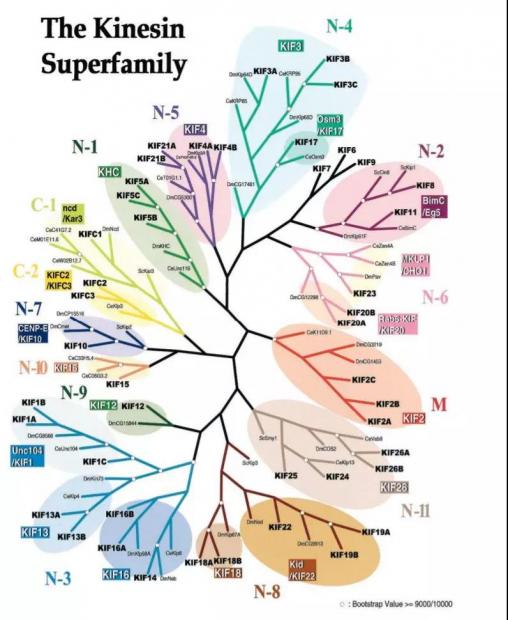
驱动蛋白家族成员众多 来源:Ref.30
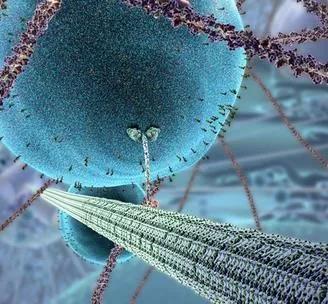
Kinesin,细胞内的“快递小哥” 来源:The Inner Life of the Cell (2006) Harvard Biology
新的征程(1985-现在)
时间荏苒,当年意气风发的小麦如今已经年逾古稀。但依然奋战在科研一线。不同于专注于阐释驱动蛋白结构和功能的罗纳德,迈克尔在包含囊泡内吞过程膜张力变化、整合素感受蛋白互作张力在内的多个领域都有涉猎,其中对于生物力学的关注是他几十年来的一条研究主线。[31-34]
在发现驱动驱动蛋白后不久,迈克尔· 希茨将实验室搬到了圣路易斯的华盛顿大学,随后又来到杜克大学医学院担任系主任。他于1990年起受聘哥伦比亚大学讲席教授。2009年,他在新加坡国立大学牵头建立了生物力学研究所并任教授。
2012年,迈克尔· 希茨、詹姆和斯·斯普迪赫和罗纳德·韦尔凭借他们对于分子动力蛋白的工作被授予拉斯克基础医学奖。
2019年5月,希茨受聘德克萨斯大学医学部韦尔奇讲席教授。
参考资料
1. Sheetz, M. P. (2012). Following nature'schallenges. Nature medicine, 18(10), 1483.
2.
3.https://
4. Sheetz, M. P., & Chan, S. I. (1972).Effect of sonication on the structure of lecithin bilayers. Biochemistry,11(24), 4573-4581.
5. Sheetz, M. P., & Chan, S. I. (1972).Proton magnetic resonance studies of whole human erythrocyte membranes.Biochemistry, 11(4), 548-555.
6. Glaser, M., Simpkins, H., Singer, S. J.,Sheetz, M., & Chan, S. I. (1970). On the interactions of lipids andproteins in the red blood cell membrane. Proceedings of the National Academy ofSciences, 65(3), 721-728.
7. Sheetz, M. P., & Singer, S. J. (1974).Biological membranes as bilayer couples. A molecular mechanism ofdrug-erythrocyte interactions. Proceedings of the National Academy of Sciences,71(11), 4457-4461.
8. Sheetz, M. P., Schindler, M., & Koppel,D. E. (1980). Lateral mobility of integral membrane proteins is increased inspherocytic erythrocytes. Nature, 285(5765), 510.
9. Huxley, H. E. (1969). The mechanism ofmuscular contraction. Science, 164(3886), 1356-1366.
10. Huxley, A. F., & Niedergerke, R. (1954).Structural changes in muscle during contraction: interference microscopy ofliving muscle fibres. Nature, 173(4412), 971.
11. Huxley, H., & Hanson, J. (1954). Changesin the cross-striations of muscle during contraction and stretch and theirstructural interpretation. Nature, 173 (441) 973
12. Spudich, J. A., Huxley, H. E., & Finch,J. T. (1972). Regulation of skeletal muscle contraction: II. Structural studiesof the interaction of the tropomyosin-troponin complex with actin. Journal ofmolecular biology, 72(3), 619-632.
13. Spudich, J. A. (2012). One path tounderstanding energy transduction in biological systems. Nature medicine,18(10), 1478.
14. Kersey, Y. M., Hepler, P. K., Palevitz, B.A., & Wessells, N. K. (1976). Polarity of actin filaments in Characeanalgae. Proceedings of the National Academy of Sciences, 73(1), 165-167.
15. Clarke, M., & Spudich, J. A. (1974).Biochemical and structural studies of actomyosin-like proteins from non-musclecells: isolation and characterization of myosin from amoebae of Dictyosteliumdiscoideum. Journal of molecular biology, 86(2), 209-222.
16. Sheetz, M. P., & Spudich, J. A. (1983).Movement of myosin-coated fluorescent beads on actin cables in vitro. Nature,303(5912), 31.
17. Vale, R. D., De Lean, A. N. D. R. E.,Lefkowitz, R. J., & Stadel, J. M. (1982). Regulation of insulin receptorsin frog erythrocytes by insulin and concanavalin A. Mol. Pharmacol, 22,619-626.
18. Vale, R. D., & Shooter, E. M. (1983).Conversion of nerve growth factor-receptor complexes to a slowly dissociating,Triton X-100 insoluble state by anti nerve growth factor antibodies.Biochemistry, 22(21), 5022-5028.
19. Brady, S. T., Lasek, R. J., & Allen, R.D. (1982). Fast axonal transport in extruded axoplasm from squid giant , 218(4577), 1129-1131.
20. Allen, R. D., Metuzals, J., Tasaki, I.,Brady, S. T., & Gilbert, S. P. (1982). Fast axonal transport in squid giantaxon. Science, 218(4577), 1127-1129.
21. Inoue, S. 1981. Video image processinggreatly enhances contrast, quality and speed in polarizationbased microscopy.J. Cell Biol. 89:346–356
22. Baraff, L. S., & Loughlin, T. R. (2000).Trends and potential interactions between pinnipeds and fisheries of NewEngland and the US West Coast. Marine Fisheries Review, 62(4), 1-39.
23. Adams, R. J., & Bray, D. (1983). Rapidtransport of foreign particles microinjected into crab axons. Nature,303(5919), 718.
24.
25. Schnapp, B. J., Vale, R. D., Sheetz, M. P.,& Reese, T. S. (1985). Single microtubules from squid axoplasm supportbidirectional movement of organelles. Cell, 40(2), 455-462.
26. Vale, R. D. (2012). How lucky can one be? Aperspective from a young scientist at the right place at the right time. Naturemedicine, 18(10), 1486.
27. Vale, R. D., Schnapp, B. J., Reese, T. S.,& Sheetz, M. P. (1985). Organelle, bead, and microtubule translocationspromoted by soluble factors from the squid giant axon. Cell, 40(3), 559-569.
28. Vale, R. D., Reese, T. S., & Sheetz, M.P. (1985). Identification of a novel force-generating protein, kinesin,involved in microtubule-based motility. Cell, 42(1), 39-50.
29. Kull, F.J., Sablin, P., Lau, R., Fletterick,R.J. and Vale, R.D. (1996) Crystal structure of the kinesin motor domainreveals a structural similarity to myosin. Nature 380: 550-555.
30. Miki, H., Setou, M., Kaneshiro, K., &Hirokawa, N. (2001). All kinesin superfamily protein, KIF, genes in mouse andhuman. Proceedings of the National Academy of Sciences, 98(13), 7004-7011.
31. Sheetz, M.P., Sable, J.E. & Dobereiner,H.G. Continuous membrane-cytoskeleton adhesion requires continuousaccommodation to lipid and cytoskeleton dynamics. Annu. Rev. Biophys. Biomol.Struct. 35, 417–434 10.1146/annurev.biophys.35.040405.102017 (2006).
32. Vogel, V. & Sheetz, M.P. Cell fateregulation by coupling mechanical cycles to biochemical signaling pathways.Curr. Opin. Cell Biol. 21, 38–46 10.1016/j. ceb.2009.01.002 (2009).
33. Ghassemi, S. et al. Cells test substraterigidity by local contractions on submicrometer pillars. Proc. Natl. Acad. Sci.USA 109, 5328–5333 10.1073/ pnas.1119886109 (2012).
34. Wolfenson, H., Yang, B., & Sheetz, M. P.(2019). Steps in mechanotransduction pathways that control cell morphology.Annual review of physiology, 81, 585-605.
文章来源于赛先生
话题:

0
推荐
财新博客版权声明:财新博客所发布文章及图片之版权属博主本人及/或相关权利人所有,未经博主及/或相关权利人单独授权,任何网站、平面媒体不得予以转载。财新网对相关媒体的网站信息内容转载授权并不包括财新博客的文章及图片。博客文章均为作者个人观点,不代表财新网的立场和观点。
 京公网安备 11010502034662号
京公网安备 11010502034662号 {kind=link}